Использование разнообразных методов для изучения процессов абсорбции позволило установить наличие в кишечнике механизма активного транспорта кальция. Об этом свидетельствуют следующие факторы:
- транспорт ионов кальция происходит против электрического градиента и против градиента концентрации;
- для его осуществления требуется обменная энергия, так как транспорт тормозится при депрессии метаболизма или при добавлении к раствору ингибиторов окислительного фосфорилирования;
- транспорт является относительно специфичным для кальция.
Этот активный переход кальция через кишечную стенку следует рассматривать как одно из звеньев гомеостатического механизма, регулирующего поступление и обмен кальция в организме.
В организм кальций поступает с растительными кормами и минеральными добавками. В растениях он связан с белками и анионами органических кислот, в добавках — с анионами карбоната или фосфата. Независимо от формы кальциевых соединений под влиянием желудочного сока большая часть введенного кальция (кроме оксалата) превращается в хлористый кальций, почти полностью диссоциирующий на ионы. Ионная форма, по-видимому, является основной формой абсорбируемого кальция в двенадцатиперстной кишке и частично в желудке. Непосредственный механизм переноса ионов Са2+ через мембраны кишечного эпителия до конца неясен. Предполагается участие в этом процессе особого кальцийсвязывающего белка (СаСБ), вырабатываемого клетками слизистой. СаСБ имеет молекулярную массу примерно 28 000, характеризуется высоким содержанием двухосновных карбоновых кислот и обладает повышенным сродством к кальцию. Он обнаружен в слизистой кишечника цыплят, крыс, собак, рыб; в почках и матке кур; в молочных железах млекопитающих. Индуцируется образование СаСБ метаболитами витамина D.
Точная функция этого белка пока не известна (белок-переносчик, белок-стимулятор диффузии или белок-передатчик). Локализация в зоне щеточной каймы клеток, по-видимому, исключает его роль как непосредственного трансмембранного переносчика кальция. Имеется, однако, и другая точка зрения.
Поскольку кальций всасывается в ионной форме, уровень его абсорбции в значительной мере определяется активностью агентов, снижающих количество ионизированного кальция в кишечнике. Анионы, осаждающие или связывающие ионы кальция (оксалат, фитат, фосфат, возможно, сульфат), при избытке могут препятствовать абсорбции кальция в кишечнике.
Оптимальное соотношение Ca:Р для абсорбции обоих элементов в кишечнике равно 1,3:1—1,5:1.
Так, у жвачных животных под влиянием соляной кислоты сычужного сока почти весь кальций и магний сычужного содержимого превращаются в форму, способную проходить через коллодийную мембрану. В то же время пропорция диализируемого кальция и магния резко снижается по ходу кишечного канала с увеличением pH среды.
Значительная часть растворимых соединений кальция в кишечнике снова превращается в труднорастворимые фосфорнокислые и углекислые соли и почти нерастворимые соединения кальция с высшими жирными кислотами — пальмитиновой, стеариновой, олеиновой (мыла).
В абсорбции этих соединений кальция важная роль принадлежит желчным кислотам — холевой и дезоксихолевой, образующим комплексные соединения с кальциевыми солями ненасыщенных жирных кислот — мицеллы, высокодиспергированные в водной среде частицы диаметром 30—100 ангстрем. Смешанные мицеллы способствуют растворению и всасыванию кальциевых солей насыщенных жирных кислот и сами проникают сквозь мембрану клеток, освобождая при этом жирные кислоты. Предполагается, что к «мицеллярному» переносу кальция в кишечнике имеют отношение и ионы Na+.
На абсорбцию кальция отрицательно влияет избыток фосфатов, жиров, ионов магния, алюминия. Благоприятствуют всасыванию витамин D3, а также вещества, снижающие pH кишечника (например, лактоза) или образующие с кальцием легкорастворимые металлохелаты.
В расчете на единицу поверхности слизистой наиболее активная абсорбция кальция у всех видов животных происходит в двенадцатиперстной кишке и верхнем отделе тощей кишки. По-видимому, чем ближе к желудку, тем в большей степени функционирует механизм переноса ионного кальция; по ходу кишечника начинает преобладать механизм «мицеллярного» переноса, предполагающий более медленное освобождение ионизированного кальция и поступление его в кровь.
Как уже отмечалось, величину истинной абсорбции кальция из корма можно определить только с учетом его эндогенных потерь, внося соответствующую поправку в показатели кажущейся абсорбции. Эндогенные потери колеблются у разных видов животных в пределах от 18 до 50 мг на 1 кг живой массы в сутки. У жвачных они больше, чем у животных с однокамерным желудком, поэтому показатели истинной абсорбции кальция (и фосфора) у взрослых жвачных в 1,5—2 раза выше, чем показатели кажущейся абсорбции.
Относительно мест абсорбции кальция у жвачных мнения противоречивы, однако многие исследователи указывают на всасывание кальция в преджелудках и сычуге. В зависимости от возраста животных и других факторов абсорбция колеблется от 10 до 50% (2, 21, 75, 92 и др.) и, по-видимому, мало зависит от содержания азота в рационе. При обычных условиях кормления этот показатель у взрослых животных составляет 10—25%. Выделением кальция со слюной и желудочным соком при этом пренебрегают, принимая во внимание его низкую концентрацию в секретах.
По мнению некоторых авторов, в тонком кишечнике овец валовое количество кальция не уменьшается, а, наоборот, возрастает в результате его эндогенной экскреции; основным же местом абсорбции являются сложный желудок и толстый отдел кишечника. В опытах на телятах с использованием неабсорбируемой метки всасывание, однако, наблюдалось во всех отделах кишечника и зависело от состава рациона. Возможно, имеются видовые и возрастные особенности абсорбции кальция и у жвачных.
Концентрация кальция в содержимом желудочно-кишечного канала у 3-месячных телят (мг на 1 г потребленного корма). Рассчитано по отношению Ca:Cr2O3 в разных отделах после убоя животных
| Отделы желудочно-кишечного канала | Состав рациона | |
| концентраты | концентраты+сено | |
| Рубец | 4,4 | 8,0 |
| Сычуг | 8,2 | 10,4 |
| Тонкий кишечник (верхняя треть) | 3,9 | 5,1 |
| Тонкий кишечник (нижняя треть) | 2,8 | 5,6 |
| Слепая кишка | 2,1 | 4,8 |
| Толстый кишечник | 1,8 | 4,5 |
| Ca в корме, мг/г | 6,7 | 7,5 |
| Ca всего в корме за день (г) | 19,0 | 20,3 |
Дальнейшее изучение этого вопроса представляет большой интерес, ибо принято считать, что рубцовый эпителий относительно непроницаем для кальция и магния.
Кальций, всасывающийся из кишечника, поступает через воротную вену в печень, где комплексные соединения его расщепляются и кальций образует какие-то новые соединения, возможно, с белками. В печени кальций, подобно другим катионам, на некоторое время задерживается, что обеспечивает его более или менее равномерное поступление в периферическую кровь. Кровь представляет собой «центральный метаболический котел», через который проходит кальций в процессе его всасывания, отложения в мягких тканях, перемещения из одной в другую и при экскреции.
Материалы, приведенные ниже, дают общее представление об основных параметрах обмена кальция в организме сельскохозяйственных животных.
Средние показатели обмена кальция у животных
| Кальций, мг в день | Теленок | Овца | Коза | Свинья |
| Живая масса, кг | 50 | 43 | 72 | 35 |
| Потребляемый | 5 800 | 3 000 | 9400 | 11 000 |
| Обмениваемый тела, мг | 33 000 | 20 000 | 3200 | 14 900 |
| Абсорбируемый | 5 300 | 800 | 1100 | 4700 |
| Эндогенный фекальный | 500 | 500 | 800 | 1450 |
| Выводимый с калом | 1 120 | 2 700 | 9100 | 7800 |
| Выделяемый с мочой | 10 | 50 | 80 | 110 |
| Откладываемый в скелете | 15 000 | 5 000 | 2300 | 13 300 |
| Извлекаемый из скелета | 10 300 | 4 750 | 2250 | 10200 |
У беременных и лактирующих животных (особенно в конце беременности и начале лактации) параметры метаболизма кальция резко меняются: увеличиваются выход его из костей, эндогенная фекальная экскреция, уровень абсорбции в кишечнике. Большое количество минеральных веществ, в том числе и кальция, выделяется в начале лактации с молозивом. Баланс кальция в этот период может быть отрицательный. Во второй половине лактации он становится положительным, усиливается отложение кальция в скелете.
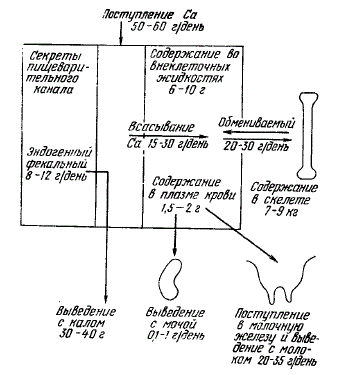
Обмен кальция у лактирующей коровы
В нормальных условиях кальций (как кормового, так и эндогенного происхождения) выводится из организма в основном через желудочно-кишечный канал.
В отличие от одновалентных ионов (К+, Na+), которые могут почти полностью абсорбироваться в кишечнике, а затем выделяться из организма без нарушения гомеостатического механизма их почечной регуляции, фильтруемый в клубочках почек кальций почти на 99% реабсорбируется в извитых канальцах, в связи с чем уровень его экскреции с мочой ограничен. Выделяющееся с мочой небольшое количество кальция связано в неионизированных фильтрующихся комплексах, по-видимому, с цитратом. При введении цитрата в кровь скорость экскреции кальция с мочой резко возрастает.
Сказанное в принципе относится к большинству лабораторных и сельскохозяйственных животных, хотя имеются видовые и возрастные различия. Так, у свиней, кроликов и кур-несушек отмечается более высокое, чем у других видов животных, выделение кальция с мочой. У кур кальциурия может колебаться от 40 до 250 мг%, в зависимости от наличия или отсутствия формирования яйца в яйцеводе. Высока концентрация кальция (до 100 мг%) в моче телят, поскольку в первые две недели жизни жвачных кальций выделяется в основном через почки, а не через кишечник.
Результаты балансовых опытов на жвачных животных систематизированы в работе Ю. К. Олля. Приведенные им материалы показывают, что в обычных условиях коровы выделяют с мочой незначительное количество кальция (0,2—1,0 г в сутки). При даче 45Са per os лактирующим коровам за 15 дней опыта с мочой выделялось 1,3—2,0% введенной дозы изотопа, а с молоком — 6,0—8,6% при среднем усвоении от 19,1 до 28% введенной дозы.