Потребности животных в сере удовлетворяются главным образом за счет серосодержащих аминокислот и частично гетероциклических соединений — биотина и тиамина. Неорганическая (сульфатная или сульфитная) сера поступает с кормами в незначительном количестве и обычно не играет большой роли в питании.
Сера абсорбируется в тонком отделе кишечника. Свободные аминокислоты, сульфатиды, тиамин, пиридоксин, биотин всасываются, не расщепляясь, а серосодержащие аминокислоты, входящие в состав белка, — после расщепления последнего. Неорганический сульфат всасывается, но в сравнительно небольшой степени.
При недостатке в рационе серы удержание в организме меченых серосодержащих соединений возрастает, при дисбалансе аминокислот — снижается.
В жидкости рубца жвачных концентрация общей серы колеблется в значительных пределах (50—500 мг/л) в зависимости от поступления ее с кормом. Сера необходима микроорганизмам для переваривания клетчатки, утилизации небелковых источников азота, для синтеза витаминов группы В.
В биохимическом отношении особенностью обмена серы у жвачных животных является способность некоторых видов рубцовых микроорганизмов (и в меньшей степени микроорганизмов, населяющих слепую кишку у моногастричных животных и птиц) утилизировать неорганическую серу и включать ее в серосодержащие аминокислоты и белки.
Возможность синтеза S-аминокислот микроорганизмами была неоднократно продемонстрирована добавлением элементарной серы или сульфата к синтетическому рациону, недостаточному по сере с частичной заменой азота протеина мочевиной.
Положительный эффект был получен также и при добавках серы к некоторым практическим рационам, содержащим часть азота в форме мочевины и включающим корма, выращенные на почвах, дефицитных по сере.
Опыты с введением жвачным per os 35S-сульфата показали, что радиоактивная сера включается в цистин и метионин микробиального белка, а после расщепления последнего всасывается и поступает в ткани. В печени при этом обнаруживается 1,8—2% введенной дозы 35S, причем более 4/5 — в виде цистеина и метионина. У овец в течение длительного времени радиоактивная сера включается в цистеиновую фракцию волокон шерсти. Таким образом, очевидно, что микроорганизмы преджелудков способны синтезировать серосодержащие аминокислоты из аммиака, углеводов и минеральной серы и что в определенных условиях сера может оказаться лимитирующим фактором. Добавки в этих условиях сульфатов или (с меньшим эффектом) элементарной серы при отношении N:S, равном 10:1, положительно влияют на рост и продуктивность животных.
Пути превращения минеральной серы в цистин и метионин микроорганизмами пищеварительного тракта иллюстрируются схемой:
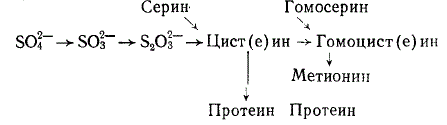
Восстановление микроорганизмами сульфатов и сульфитов до сульфидов и включение сульфидной серы в аминокислоты отмечается также в пищеварительном тракте кроликов (особенно при свободной копрофагии), птиц и свиней.
Распределение радиоактивной серы в печени разных видов животных при введении per os 35S-сульфата
| Показатели | Свинья | Кролики нормальные | Кролики лишенные копрофагии | Овца |
| 35S в печени, % дозы | 0,38 | 0,32 | 0,06 | 1,85 |
| В том числе: | ||||
| 35S-сульфаты | 98,6 | 29,3 | 85,0 | 18,4 |
| 35S-аминокислоты (цистин + метионин) | 1,4 | 70,7 | 15,0 | 83,6 |
При отсутствии копрофагии 85% всей радиоактивной серы, обнаруженной в печени кроликов, было представлено сульфатом (у контрольных животных лишь 29%). Это свидетельствует о более высокой трансформации минеральной серы в органическую у животных без ограничения копрофагии. Опыты с внутривенным введением 35S-сульфата подтвердили, что минеральная сера включается в цистеин, метионин и витамины группы В только через микрофлору пищеварительного тракта.
Частичная абсорбция 35S-соединений наблюдается в рубце жвачных и желудке моногастричных животных, однако основной процесс всасывания происходит в кишечнике. Всасывание и усвоение серы аминокислот определяется уровнем протеинового и энергетического питания животных и осуществляется с помощью механизмов активного транспорта. Сера сульфатов и. сульфитов, поступающая с кормом, абсорбируется, по-видимому, простой диффузией. В крови птиц радиоактивная сера обнаруживается уже через несколько минут после перорального введения 35S-метионина с максимумом концентрации на первом-втором часу. В период от 6 до 24 ч радиоактивность остается постоянной и довольно высокой.
Распределение абсорбированной радиоактивной серы по органам и тканям в общем однотипно для всех животных и всех исследованных 35S-соединений, однако обнаруживаются и некоторые различия.
Распределение радиоактивной серы по органам и тканям цыплят при пероральном введении им различных 35S-соединений. В процентах от общей радиоактивности, обнаруженной в тушках цыплят.
| Органы и ткани | Введенные 35S-соединения | ||
| метионин | цистеин | сульфат | |
| Мышцы + кости | 52,3 | 38,72 | 56,18 |
| Печень | 8,65 | 4,31 | 4,06 |
| Кожа | 7,93 | 7,44 | 3,37 |
| Перья | 7,51 | 9,41 | 1,45 |
| Мышечный желудок | 6,81 | 21,33 | 15,17 |
| Кишечник | 5,41 | 5,99 | 5,62 |
| Кровь | 2,58 | 2,28 | 4,31 |
| Прочие ткани | 8,72 | 8,52 | 8,84 |
| Всего | 100 | 100 | 100 |
Наиболее высокая относительная активность (в % к средней) наблюдается при введении 35S-метионина в мышцах, печени и коже; при введении 35S-цистеина — в мышечном желудке, перьях, кишечнике; при введении сульфата — в мышечном желудке, костях.
При пероральном введении курам 35S-тиамина основная часть задержанного радиоизотопа обнаруживается в стенках кишечника, мышцах, печени, почках; заметная активность наблюдается в коже, перьях и костях.
Показано, что радиоактивная сера 35S-метионина быстрее всего включается в глютатион и белковую фракцию крови, печени и мышц и медленнее — в те же фракции других органов. Сульфидная фракция медленнее фиксирует 35S, чем белковая и глютатионная.
Большая часть (до 90%) парентерально введенного животным 35S-сульфата выводится с мочой в неизменном виде или в составе эфиров. В крови, печени, мозге концентрация быстро падает; вместе с тем наблюдается заметное поглощение радиоактивной серы костями, костным мозгом и хрящами, с последующим медленным замещением 35S стабильной серой. В костях, коже, хряще 35S обнаруживается в составе хондроитин-сульфата, в шерсти овец и перьях птицы — в составе цистина, в сперме животных и кутикуле мышечного желудка — в составе неидентифицированных протеинов.
Для формирования шерсти и пера сера сульфата используется, по-видимому, в той же степени, что и сера метионина, но меньше, чем сера цистина.
В белке и желтке снесенных яиц 35S содержится в форме сульфата, 35S-цистина и в очень небольшом количестве в виде 35S-метионина. В молоке радиоактивная сера, также появляется очень быстро при введении любых растворимых 35S-соединений. Сера молока (ее около 0,03%) представлена серосодержащими аминокислотами, лактоальбумином и лактоглобулином, минеральными сульфатами (0,15 мг%) и другими свободными серосодержащими соединениями.
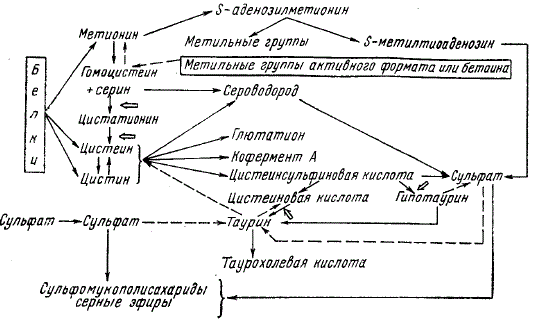
Схема обмена серы в организме нежвачных животных. Стрелками указаны точки приложения пиридоксальфосфата
Общая схема метаболизма соединений серы в организме животных приведена на рисунке. Практически единственным источником образования серосодержащих соединений в теле служат метионин и цистеин, хотя в процессы синтеза включается также небольшое количество сульфата. Сера метионина используется для построения цистеина с участием серина (процесс необратимый) через стадии гомоцистеина и цистатионина. Гомоцистеин наряду с превращением в процессе деструктивного распада в цистеин может подвергаться метилированию за счет «активного формиата», отщепляемого от глицина, серина и других соединений, а также за счет бетаина. В этом случае происходит ресинтез метионина.
При превращении цистеина в цистин образуется цистеинсульфиновая кислота, осуществляющая связь между обменом аминокислот и обменом углеводов в организме. Эта кислота может подвергнуться декарбоксилированию (с образованием гипотаурина, затем таурина), трансаминированию или окислению. Цистеиновая кислота также декарбоксилируется с образованием таурина.
При окислении цистеина, цистина и метионина в организме высших животных в качестве конечного продукта метаболизма образуется сульфат. Метаболизм сульфата у птиц несколько отличается от метаболизма у млекопитающих животных. В то время, как млекопитающие, по-видимому, удовлетворяют свои физиологические потребности в ионах SO24- за счет сульфатов, образующихся в процессе метаболизма серосодержащих аминокислот, у цыплят был обнаружен небольшой ростовой эффект от прибавления сульфата к синтетическому рациону (исключая описанные выше ситуации для жвачных). Поскольку неорганическая сера не использовалась для синтеза аминокислот в количествах, достаточных для объяснения ростового эффекта, возникло предположение, что этот эффект отражает потребности цыплят именно в сульфате.
Наличие в печени и белке яиц 35S-аминокислот после инъекции курам 35S-сульфата некоторые авторы объясняют не образованием аминокислот из сульфата в процессе межуточного метаболизма, а экскрецией меченых соединений (особенно таурина) в просвет кишечника с последующим синтезом 35S-метионина кишечной микрофлорой. Таурин у птиц, по-видимому, нельзя рассматривать как конечный продукт обмена аминокислот. В тканях птиц таурин содержится в большей концентрации, чем у млекопитающих, и выполняет другие функции, помимо его участия в синтезе таурохолевой кислоты желчи. Одна из функций таурина, возможно, состоит в том, что он является промежуточным продуктом в превращении неорганической серы в серу аминокислот.
В разных тканях организма цыплят обнаружены ферменты, необходимые для синтеза таурина из сульфата. Однако дальнейшая судьба таурина изучена недостаточно. При инъекции таурина цыплятам высокая радиоактивность отмечалась в небелковых экстрактах мягких тканей, а в печени и почках, кроме того, в тауринбелковом комплексе. В метиониновой или цистеиновой фракции органов радиоактивная сера не обнаружилась. Эти данные показывают, что у яичных цыплят превращение серы таурина в серу аминокислот незначительно.
Куры-несушки, вероятно, могут синтезировать небольшое количество цистина (но не метионина) в процессе тканевого метаболизма из абсорбируемой в кишечнике неорганической серы (через таурин → цистеиновую кислоту?). Однако включение сульфатной серы в аминокислоты слишком мало, чтобы иметь значение в протеиновом питании несущейся птицы. В рационах бройлеров можно использовать неорганическую серу для частичного восполнения потребности в цистине. Наряду с этим сульфату приписывается значение самостоятельного ростового фактора, а также подчеркивается его участие в детоксикационных процессах в организме бройлеров.
Основная часть продуктов обмена серы (сульфаты в свободном или эфиросвязанном состоянии, таурин, тиосульфат, сульфоционат и др.) выделяется из организма с мочой. В моче сера содержится в трех формах: минеральной, эфиросвязанной и нейтральной. Указанные фракции составляют соответственно 80—85%, 5—8 и 10—14% от общей серы мочи. Минеральная (сульфатная) сера происходит частично из абсорбированных, но не усвоенных сульфатов корма, а главным образом из окисленных аминокислот.
Ионы SO24- экскретируются с эквивалентным количеством катионов Na+, К+ и NH4+ При неизменном уровне протеина в рационе соотношение азота и серы в моче достаточно постоянно. Оно уменьшается при голодании или недостатке протеина и возрастает при увеличении уровня белка в рационе.
Сера эфиров представлена соединениями серной кислоты либо с фенолом или крезолом (производными соответственно фенилаланина и тирозина), либо с индоксилом и скатоксилом (производными триптофана). Эти вещества образуются в процессе кишечного гниения под действием бактерий. Связывание их в печени и частично в тонком кишечнике и почках рассматривается как детоксикационный механизм.
Нейтральная сера — это сера аминокислот (в основном цистина), таурина, меркаптана, тиамина, биотина, урохрома. Увеличение в моче количества нейтральной серы служит показателем усиленного распада эндогенного белка.