Одним из первых доказательств сложности гена явилось обнаружение явления множественного аллелизма.
Оказалось, что в результате мутирования ген может находиться более чем в двух различных состояниях. Серии множественных аллелей найдены для разных локусов у самых различных организмов. Известны серии, включающие десятки аллелей. Так, число аллелей, составляющих серию, определяющую клеточные антигены у крупного рогатого скота, достигает более 100. Обнаружение множественного аллелизма свидетельствует о большей функциональной лабильности гена, чем это думали раньше. Стало ясно, что ген может находиться не только в двух крайних альтернативных состояниях — доминантном и рецессивном. Теперь допускается существование нескольких доминантных и нескольких рецессивных аллелей одного гена. В результате изучения множественного аллелизма было установлено, что, мутируя в различные состояния, ген остается при этом делимой единицей, занимающей определенный локус в хромосоме, и его аллели представляют собой лишь изменения одного и того же локуса.
Представление об аллельном состоянии наследственного фактора прямо вытекало из работ Г. Менделя, из открытого им явления доминантного и рецессивного состояния гена. В. Бэтсон и Т. Морган придали новый смысл понятию аллелизма (аллеломорфизма).
В 1929—1930 гг. в нашей стране в работах А. С. Серебровского и его молодых сотрудников — Н. П. Дубинина, Б. Н. Сидорова др. был впервые поставлен вопрос о функциональной сложности авторы исследовали серию множественных аллелей у дрозофилы локуса scute, локализованного на нулевой морганиде половой хромосомы. Мутации scl, sc2, sc3 и другие мутации этого локуса обусловливают редукцию разных щетинок на теле мухи. Было проведено тщательное изучение фенотипических эффектов, производимых мутациями этого локуса при их комбинировании. При скрещивании особей, гомозиготных по тем или иным мутантным аллелям, например sc1//sc1 x sc2// выявилась интересная картина: у гетерозигот — как правило, отсутствовали лишь те щетинки, которые были редуцированы у обеих гомозигот sc1/sc1 и sc2/sc2.
Так, например, если одна мутация sc1 вызывала редукцию щетинок ABC, а другая sc2 — редукцию щетинок BCD, то у гетерозиготы от sc1/sc2 отсутствовали щетинки В и С, а щетинки А и D имелись. Создавалось впечатление, что в данном случае речь идет о частичной гетерозиготности, когда части мутантных аллелей, которые обусловливают Знаковый фенотипический эффект, оказываются в гомозиготном состоянии. Всего было исследовано 13 различных мутаций в локусе и при их сочетании наблюдалась одна и та же закономерность.
Если эту закономерность представить графически, то образуется как бы лестница, ступенями которой служат различные аллели sc:
sc1 — ABC
sc2 — BCD
sc3 — CDE
Поэтому описанное явление получило название ступенчатого аллелизма. Согласно гипотезе авторов, ген представлялся как основа — базигена, составленная из частей — трансгенов. Вместе с тем авторы совсем не изучали структурную делимость гена посредством кроссинговера.
Представление о гене как единице, далее не делимой кроссинговером, подразумевало, что при гаметогенезе у компаундов, т. е. зигот, несущих две рецессивные аллели одной серии (a1/a2) могут образовываться гаметы только двух типов а1 и а2. При возвратном скрещивании таких особей с любой из родительских форм возможно появление только мутантных форм: a1/a2 x a1/a1 → a1/a1 : a2/a1.
Действительно, это и наблюдается при исследовании ограниченной выборки потомков от возвратного скрещивания. Однако если выборку увеличить, например, до 100 тыс. и более особей, то в ней окажутся и потомки дикого типа. Такие особи могли появиться только в результате кроссинговера, приводящего к обмену частями между аллелями а1 и а2 одного и того же мутантного гена.
Это явление было открыто в конце тридцатых — начале сороковых годов при изучении ряда генов у дрозофилы Е. Льюисом, М. Грином, К. Оливером и др. Существование такого явления противоречило представлению о гене как о единице, далее не делимой при перекресте. Однако от этих традиций трудно было сразу отказаться, и об аллелях, дающих при кроссинговере фенотип дикого типа, стали говорить, как о псевдоаллелях.
Первоначально полагали, что псевдоаллелизм встречается в виде редких исключений, но с увеличением разрешающей способности генетического анализа, когда число анализируемых особей в исследуемых выборках резко возросло, становилось все более ясным, что данное явление распространено весьма широко. Оно было продемонстрировано на разнообразных организмах: аспергилле, нейроспоре, дрожжах, хлопчатнике, кукурузе, шелкопряде, дрозофиле, голубях, мышах и многих других объектах. В скором времени большинство исследователей пришло к выводу, что псевдоаллелизм представляет собой скорее правило, чем исключение.
Одновременно с накоплением примеров псевдоаллелизма создавалось представление о тесном функциональном родстве псевдоаллелей. Так, диплоиды типа a1/a2, a2/a3, a1/a3 всегда имели мутантный фенотип.
Период исследования псевдоаллелизма был переходом к формированию современной теории гена, руководствующейся как рекомбинационным, так и функциональным критериями для понимания аллелизма.
Аллелями называются различные состояния одного гена. Соответственно аллельными мутациями называются мутации, затрагивающие один и тот же ген. Исходя из того, что ген, согласно новейшим открытиям, представляет собой сложную линейную структуру, мутации могут затрагивать различные его участки.
Естественным будет вопрос: как же практически определить аллельны или нет две какие-либо независимо возникшие мутации? Каков же критерий аллелизма? Впервые на эти вопросы ответил Т. Морган. Он рассматривал два критерия аллелизма для рецессивных мутаций: функциональный и рекомбинационный.
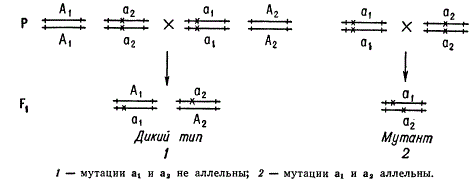
Функциональный тест на аллелизм, предложенный Морганом
Функциональный критерий основывается на том, что при скрещивании двух мутантов, несущих изменения разных генов, возникает гибрид первого поколения — дигетерозигота, имеющая дикий фенотип в силу доминирования нормальных, аллелей каждого из генов. В таком случае принято считать, что исследуемые рецессивные мутации комплементарны друг другу. В то же время, бели скрещиваемые мутанты несут в компаунде аллельные мутации, то дикий тип не появляется, так как обе аллели, одного и того же гена, находящиеся в гомологичных хромосомах, несут мутационные изменения т. е. эти мутации некомплементарны.
Этот критерий особенно важен в свете того, что могут быть фенотипически сходны мутанты, несущие изменения разных генов. Например, у дрозофилы мутация в любом из следующих генов: v (vermilion), cd (Cardinal), st (scarlet), p (purple) выражается в исчезновении бурого глазного пигмента. Мутации, находящиеся в разных генах, способны рекомбинировать между собой. В то же время при исследовании гибрида, несущего аллельные мутации, орган и его сотрудники рекомбинации не обнаруживали (как теперь понимаем, в силу ограниченности размера выборки) исследователи школы Моргана считали мутации аллельными, если соблюдались функциональный (гетерозигота — мутант) и рекомбинационный (мутации не рекомбинируют) критерии.
Абсолютизация рекомбинационного критерия привела к тому, что обнаружение кроссинговера между некомплементарными, по Моргану — аллельными, мутациями было воспринято почти как крушение теории гена. Трудно было сразу отказаться от представления о существовании какой-то далее не делимой единицы мутации, рекомбинации и функции. И когда убедились, что ген является делимой единицей, то стали искать субгены, псевдоаллели и другие элементарные единицы гена.
Исходя из таких представлений были сделаны попытки модернизировать моргановский функциональный критерий аллелизма. В частности Е. Льюис предложил так называемый цис-транс-тест на аллелизм.
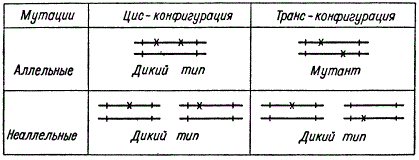
Цис-транс-тест на аллелизм
Смысл этого теста сводится к тому, что при скрещивании двух особей, несущих мутации в негомологичных участках хромосомы, возникает зигота с транс-конфигурацией этих мутаций a1/a2. Если мутации комплементарны, т. е. появляется гибрид дикого типа, то мутации относят к разным геном. Если же гибрид оказывается мутантным, то обе мутации относят к одному гену, т. е. считают их аллельными. При скрещивании двух особей, одна из которых несет два генотипических изменения, а другая представляет собой дикий тип, образуется зигота с реконфигурацией мутаций. Гибрид дикого типа возникает и тогда, когда обе мутации затрагивают один ген и когда оказываются мутантными два разных гена. Таким образом, цис-транс-тест сводится фактически к транс-тесту, т. е. к функциональному критерию аллелизма, предложенному еще Морганом.
Рассмотрим пример, который позволит более ясно понять, в чем специфика критерия комплементарности в сравнении с рекомбинационным критерием для доказательства аллельных отношений. В качестве примера могут служить опыты Гв. Понтекорво по анализу мутантов у гриба Aspergillus, нуждающихся для своего роста в аденине.
В различных опытах было получено 50 мутаций независимого происхождения, определяющих потребность в аденине. Каждый S мутант получил номер от ad1 до ad50. Необходимо было решить, являются ли эти мутации аллельными или относятся к разным генам. Для этого мутанты были испытаны в скрещиваниях. Каждую пару мутаций поочередно объединяли в гетерозиготу в транс-положении: если гетерозигота обнаруживала мутантный фенотип, делали заключение, что эти мутации — изменения одного гена; в противоположном случае соответствующие мутации относили к разным генам.
В результате такого анализа все 50 мутаций распределились по шести группам, которые получили название по какому-нибудь одному мутанту: ad1, ad3, ad8, ad9, ad20 и ad23. Гетерозиготы по мутациям внутри каждой группы давали мутантный фенотип, гетерозиготы от скрещивания мутантов из разных групп имели нормальный фенотип. Следовательно, можно было полагать, что мутации внутри каждой такой группы являются аллелями одного гена.
Затем путем учета рекомбинаций была произведена локализация мутаций всех шести групп. При этом обнаружилось, что каждая из групп заняла свой особый локус в I и II группах сцепления. Эти локусы оказались расположенными на разных расстояниях друг от друга. Однако два локуса, а именно ad1 и ad3, во II группе сцепления расположились очень близко. Возник вопрос: являются ли они самостоятельными локусами или объединяются в один локус?
С помощью критерия комплементарности было показано, что они действительно являются двумя отдельными генами, хотя и близко расположенными, находящимися на расстоянии 0,5% перекреста. В настоящее время очевидно, что ген не представляет собой единицы, далее не делимой при кроссинговере, и не представляет пробой единицы мутирования. Однако ген как единица наследственной информации остается функционально неделимым.
Итак, критерий комплементарности вместе с рекомбинационным критерием составляют ту основу, на которой базируется в настоящее время генетический анализ при установлении аллельных отношений. Независимо возникающих мутаций, сходных по своему проявлению.