Полиплоиды, возникающие на основе умножения разных геномов, называются аллополиплоидами.
Аллополиплоиды образуются на основе скрещиваний различных видов, и объединяют различные геномы. В 1927 г. М. С. Навашин предложил называть полиплоиды, возникающие в результате гибридизации и имеющие сумму наборов хромосом обеих родительских форм, амфидиплоидами. Так, например, если у межвидового гибрида совмещаются геномы А и В, то полученный от него амфигаплоид (аллодиплоид) будет АВ, гибрид с удвоенными геномами, например ААВВ, будет амфидиплоидом (аллотетраплоидом), а ААААВВВВ — аллооктоплоидом.
Полуторный набор геномов разных видов (АВВ или ААВ) по предложению М. А. Розановой называют сесквиполиплоидом.
Аллополиплоидию иначе называют гибридной полиплоидией. Она возможна у отдаленных гибридов. Гибриды от скрещивания разных видов и родов, которые обладают разными наборами хромосом, называют отдаленными гибридами. При скрещивании пшеницы и ржи возникает отдаленный ржано-пшеничный гибрид, в котором совмещаются гаплоидные наборы хромосом ржи и пшеницы. Хромосомные наборы аллополиплоида различны не только по числу хромосом, но и по их генетическому составу.
Особенности мейоза у аллополиплоидов
Часто гибрид F1 от скрещивания двух разных видов оказывается бесплодным (например, гибриды ржи с пшеницей, редьки с капустой и др.).
Рассмотрим причины этого явления. Допустим, совмещаются геномы пшеницы T и ржи S, тогда ржано-пшеничный гибрид будет нести два разных генома —(TS). При удвоении набора хромосом у такого гибрида возникнет амфидиплоид TTSS который по существу является двойным диплоидом, т. е. аллоотетраплоидом. Рожь привносит в зиготу гибрида свой геном, состоящий из семи хромосом, пшеница — геном Т, также представленный семью хромосомами. Такой отдаленный гибрид в F1 имеет в соматических клетках общее число хромосом в процессе нормального развития половых клеток: в профазе мейоза должна происходить попарная конъюгация гомологичных хромосом. Но так как в наборе хромосом пшеницы нет гомологов для хромосом ржи, то каждая из хромосом ведет себя в мейозе самостоятельно, как унивалент. В клетках указанного гибрида в мейозе можно насчитать 14 унивалентов. В анафазе редукционного деления они будут беспорядочно распределяться к полюсам, и в силу этого образуются гаметы с различным числом хромосом от 0 до 14. У такого гибрида не происходит нормального развития гамет, и поэтому он оказывается стерильным, а при частичной гомологии хромосом — с пониженной плодовитостью.
У этого гибрида некоторая часть гамет будет нести 14 хромосом: 7Т+7S, эти гаметы называются нередуцированными. При объединении в процессе оплодотворения нередуцированных гамет образуется зигота с удвоенным набором хромосом обоих видов — амфидиплойд (аллотетраплоид). Аллотетраплоид имеет два набора хромосом ржи: 7S + 7S и два набора пшеницы: 7Т + 7Т, т. е. всего (2n = 28) Он оказывается фертильным, так как в мейозе каждая хромосома имеет партнера, с которым и конъюгирует. При этом образуется 7 бивалентов ржи и 7 бивалентов пшеницы. В анафазе редукционного деления члены этих бивалентов нормально расходятся к полюсам, и образуются гаметы с числом хромосом 7Т + 7S. Эти диплоидные гаметы, содержащие разные наборы хромосом, оказываются вполне нормальными и поэтому при оплодотворении вновь воспроизводят гибридный организм с двумя диплоидными наборами хромосом от разных видов.
Если диплойдная гамета одного вида, например 7А + 7А, соединяется при оплодотворении с нормальной — гаплоидной гаметой другого вида 7В, то образуется аллотриплойд 7А + 7А + 7В. Такой гибрид оказывается стерильным. Стерильность вызывается тем, что двойной набор хромосом вида А образует в мейозе биваленты, а одинарные хромосомы вида В останутся унивалентами, которые неправильно распределяются к полюсам, вследствие чего образуются неполноценные несбалансированные гаметы.
Получение плодовитых аллополиплоидов
Получение амфидиплоидов открыло возможности синтеза новых константных форм путем гибридизации и удвоения у гибридов числа хромосом.
Надо сказать, что большой вклад в, эту область исследований внесли советские генетики Г. Д. Карпеченко, М. С. Фавашин и Б. Л. Астауров. Впервые Г. Д. Карпеченко и М. С. Наташин получили амфидиплоиды у растений, а Б. Л. Астауров — у шелкопрядов при скрещивании Bombyx mori X В. mandarina.
Классическим примером синтеза новой формы является создание межродового плодовитого гибрида от скрещивания редьки (Raphaels sativus) с капустой (brassica oleracea) полученного Г. Д. Карпеченко в начале 20-х годов. Оба эти вида, относящиеся к разным годам, имеют диплоидное число хромосом, равное 18. При скрещивании редьки с капустой было получено мощное гибридное растение. Клетки его имели диплоидное число хромосом также 18, из них 9 хромосом редьки (R) и 9 капусты (В). Гибрид обильно но не завязывал семян, так как редукционное деление у него протекало ненормально. Гаметы, образующиеся у этого гибрида, Сказываются с нарушенным числом хромосом (от 0 до 18) и нежизнеспособными.
Но в отдельных как женских, так и мужских половых клетках гибрида встречались наборы хромосом обоих видов: 9R + 9В (нередуцированные гаметы). От слияния таких диплоидных гамет образовалось семя, из которого выросло растение межродового плодовитого аллотетраплоида (9R + 9R) + (9В + 9В). Такой гибрид совмещал некоторые признаки редьки и капусты и был плодовит и константен в поколениях. В его соматических клетках имелось 36. хромосом, из которых 18 — редечных и 18 — капустных. Эта новая форма, синтезированная на основе сочетания геномов двух родов, была названа рафанобрассикой (Raphanobrassica), или редечно-капустным гибридом.
Представлены плоды и хромосомные наборы рафанобрассики. У этой формы стручок оказывается комбинированным: верхняя часть от редьки, а основание стручка типа капусты. Оригинал этого аллотетраплоида хранится на кафедре генетики Ленинградского университета.
В гаметогенезе у рафанобрассики иногда могут образовываться различные гаметы — как диплоидные (9R + 9В), так и тетраплоидные (18R + 18B). Если эти гаметы соединятся с нормальными гаметами одного из исходных видов, например, редьки (9R), или с диплоидными (9R + 9В), или с тетраплоидными гаметами рафанобрассики (18R + 18B), то могут возникать формы с разным числом наборов хромосом в соматических клетках: тетраплоиды (9R + 9R) + (9В + 9В), пентаплоиды (18R + 18В) + 9R, октоплоиды (18R + 18В) + (18R + 18В).
Изображен плод растения с другим числом хромосом, а именно: 24R + 27В. Это растение развилось из зиготы, образовавшейся при слиянии диплоидной гаметы и тетраплоидной, утратившей 3 хромосомы редьки. Однако с явлением изменения числа отдельных хромосом в наборе мы познакомимся ниже.
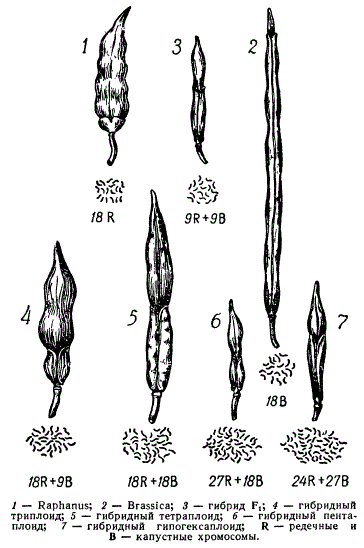
Плоды и хромосомные наборы рафанобрассики и их гибридов
На примере рафанобрассики можно видеть и влияние соотношения геномов на проявление признаков у аллополиплоидов. При равенстве геномов 18 R + 18В у рафанобрассики стручок наполовину является редечным, наполовину капустным. Аллотриплоид 9R + 9R + 9B имеет преобладание геномов редьки, и верхняя большая часть стручка оказывается типа редьки, нижняя меньшая — типа капусты. Преобладание хромосом капусты (В) обусловливает развитие большей части стручка по типу капустного. Относительно морфологических признаков гибрида Г. Д. Карпеченко в своей монографии писал: «Морфологические особенности гибридов определяются полной мере соотношением числа редечных и капустных хромосом у них и общим числом последних. Значительное накопление хромосом влияет угнетающе на развитие растения. Параллельно с увеличением числа хромосом у гибридов идет и увеличение размеров их клеток».
Аллополиплоиды были получены также при скрещивании двух видов табака — Nicotiana tabacum (2n = 48) и N. glutinosa (2n = 24). Константный амфидиплоид (аллогексаплоид) содержал 2n = 72 хромосомы. В дальнейшем межвидовые и межродовые гибриды на основе аллополиплоидии были получены у многих растений. К настоящему времени общее число аллополиплоидов, изученных в разных семействах растительного царства, составляет несколько сотен.
На основе механизма аллополиплоидии удается синтезировать новые виды. Установлено, что пшеницы новосветский длинноволокнистый хлопчатник, ягодные и плодовые культуры и некоторые другие сельскохозяйственные растения произошли именно этим путем.
Изучение аллополиплоидии на культурных растениях имеет огромное значение для создания новых форм. У нас в стране эти исследования велись О. Н. Сорокиной, В. Е. Писаревым, Н. В. Цициным, А. И. Державиным, Г. К. Мейстером, В. А. Хижняком, А. Р. Жебраком и др. В результате этих работ получены аллополиплоиды (амфидиплоиды) от гибридов между пшеницей и рожью, пшеницей и эгилопсом, пшеницей и пыреем и т. д.
А. Р. Жебраком на основе аллополиплоидии получены новые среди рода Triticum формы пшеницы:
- Т. durum (2n = 28) X Т. monococcum (2n =14) дают амфидиплоид 2n = 42 хромосомы (аллогексаплоид);
- Т. durum (2n = 28) X Т. Timopheevi (2n = 28) дают амфидиплоид 2n = 56 хромосом (аллооктоплоид);
- Т. polonicum (2n = 28) X Т. durum (2n = 28) дают амфидиплоид 2n = 56 хромосом (аллооктоплоид);
- Т. aestivum (2n = 42) X Т. Timopheevi (2n = 28) дают амфидиплоид 2n = 70 хромосом (аллодекаплоид);
- Т. Timopheevi (2n = 28) X Т. aestivum (2n = 42) дают амфидиплоид 2n = 70 хромосом (аллодекаплоид).
Все эти гибриды в диплоидном состоянии являются стерильными. При удвоении числа хромосом восстанавливается парность геномов у аллополиплоидов и гибриды становятся плодовитыми, сочетая в себе признаки и свойства скрещиваемых видов. Эти синтетические новые формы являются источником наследственной изменчивости для формирования организмов с новыми свойствами. Причем для каждого рода или вида имеется свой оптимальный уровень полиплоидии в отношении проявления как адаптивных, так и ценных в хозяйственном отношении признаков. Так, например, для мягких пшениц оптимален гексаплоидный уровень, для хлопчатника и картофеля — тетраплоидный, для земляники — октоплоидный, кукурузы, риса и тыквенных — диплоидный.
Возникновение аллополиплоидов как в естественных, так и в искусственных условиях в принципе ничем не отличается от возникновения автополиплоидов. Они могут быть митотического и мейотического происхождения. Аллоплоиды можно получить, во-первых, путем воздействия на ход митоза в точках роста отдаленного гибрида, во-вторых, путем воздействия на ход мейоза гибрида, стимулируя образование нередуцированных гамет. Митотическая полиплоидизация возможна при делении зиготы, а также и на последующих стадиях развития гибридного растения. При мейотической аллополиплоидии аллоплоиды возникают в результате слияния нередуцированных гамет при самоопылении отдаленного гибрида или при скрещивании его с одним из родительских видов.
Аллополиплоиды могут быть получены от скрещивания двух, трех и более видов. Так, например, от скрещивания двух видов А и В получается стерильный гибрид.
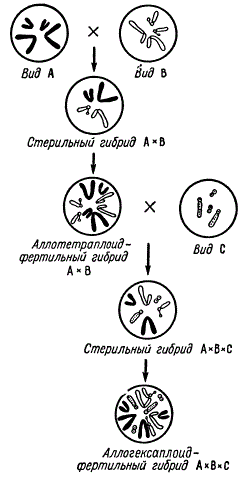
Схема получения сложных фертильных аллополиплоидов
Амфидиплоид этого гибрида будет плодовитым. Теперь его можно скрещивать еще с третьим видом — С. Тройной гибрид оказывается стерильным, так как хромосомы всех видов (А, В и С) не имеют гомологов и остаются в мейозе унивалентными. Но если удвоить набор хромосом у тройного гибрида, то он восстанавливает плодовитость.
Род пшениц представлен сложным аллоплоидным рядом. Группа мягких пшениц имеет по крайней мере три разных генома (А, В, D), каждый по семи хромосом. Хромосомы разных геномов в основном негомологичны, но среди них имеется частичная гомологичность.
Поэтому иногда даже в пределах гаплоидного набора разных геномов происходит конъюгация и образуются биваленты. В процессе эволюции изменение наборов хромосом и обмен сегментами между негомологичными хромосомами, относящимися к разным геномам, может идти одновременно. Это приводит к тому, что в мейозе аллополиплоидов могут сказываться сложные отношения между хромосомами разных геномов, обусловливающие их частичную стерильность.
Для понимания мейоза у аллополиплоидов необходимо иметь представление о явлениях, называемых автосиндезом и аллосиндезом.
Автосиндезом, или самоконъютацией, называют процесс конъюгации в профазе мейоза хромосом, происходящих от одного из родительских видов, участвующих в образовании аллополиплоида. Например, в кариотипе ААВВ хромосомы генома А конъюгируют с хромосомами А, а генома В — с В. Аллосиндезом называют конъюгацию в мейозе хромосом, происходящих от разных исходных видов. Часть хромосом вида А коньюгирует с хромосомами вида В. Наличие аллосиндеза указывает раз на то, что хромосомные наборы эта совмещенные в аллополиплоиде, имели в прошлом сходные геномы. Подобные явления служат доказательством общности происхождения данных видов.
Объединение геномов разных видов в одном растении создает высокую гетерогенность аллополиплоида, часто вызывая гетерозис. Благодаря этому в процессе естественного отбора и селекции аллополиплоидные виды завоевывают место в природе и приобретают учение в сельскохозяйственном производстве.
Совместимость и несовместимость геномов с цитоплазмой
Из того, что сказано о полиплоидах, следует, что плодовитость последних определяется характером конъюгации хромосом в мейозе, которая зависит не только от структуры гомологичных хромосом, но и от специальных генов, определяющих синапсис.
Так, например, у мягкой пшеницы Т. aestivum, имеющей 2n = 6х = 42 хромосомы, контроль формирования бивалентов, по данным Р. Райли, осуществляется генами, локализованными в V хромосоме.
Однако было бы глубокой ошибкой представлять дело так, что все, что окружает хромосомы, — цитоплазма и ее компоненты, — является нейтральной средой. Хромосомы находятся в интимной и сложной связи с цитоплазмой. Клетки функционируют как единая система и поэтому генетические различия в плодовитости полиплоидов могут определяться совместимостью или несовместимостью геномов с цитоплазмой.
Роль цитоплазмы в гаметогенезе у аллополиплоидов можно показать на примере рафанобрассики. Рафанобрассика является аллотетраплоидом с равным количеством редечных и капустных хромосом (18R + 18В). Цитоплазма этой формы принадлежит редьке, так как гибрид получен от опыления пыльцой капусты цветка редьки. Для выяснения роли цитоплазмы Г. Д. Карпеченко произвел обратные скрещивания с обеими родительскими формами: рафанобрассика X редька и рафанобрассика X капуста. При таких скрещиваниях цитоплазма сохранялась редечной, соотношение же хромосом менялось.
В первом скрещивании аллотриплоид рафанобрассика X редька имел в мейозе 9 бивалентов из редечных хромосом и 9 унивалентов хромосом капусты: (9R + 9R)+9B. Во втором скрещивании рафанобрассика X капуста было обратное соотношение хромосом: (9В + 9В) + 9R. Следовательно, оба триплоида должны иметь одинаковый ход мейоза и образование гамет. Но в первом случае 9 редечных бивалентов находятся в своей «редечной» цитоплазме, а во втором — 9 капустных бивалентов в чужой «редечной» цитоплазме.
Оказалось, что в случае второго беккросса нормальных гамет не образуется, и этот аллотриплоид полностью стерилен. В случае же первого беккросса, когда редечные хромосомы находятся в своей цитоплазме, растение является частично плодовитым. Мейоз внешне протекает сходно в обоих случаях, но в одном случае гаметы полностью гибнут, а в другом — нет.
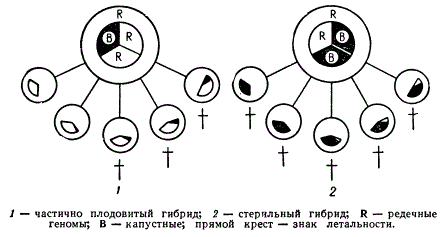
Влияние совместимости цитоплазмы Raphanus и геномов Raphanus и Brassica на фертильность аллотриплоида Raphanobrassica
Таким образом, аллотриплоиды с одинаковой цитоплазмой, но с разными наборами хромосом отличаются, так как цитоплазма в гаметогенезе у аллополиплоидов играет существенную роль.
Доказательством того, что хромосомы разных видов имеют свои физиологические особенности и ведут себя в гибридных клетках разным образом, может служить следующий опыт, проведенный недавно Н. А. Лебедевой. Она изучила реакцию хромосом на охлаждение у ряда видов картофеля: Solanum tuberosum и S. punae и др. При охлаждении ростков до + 2° хромосомы соматических клеток S. tuberosum заметно не изменяют своей формы хромосомы S. punae при тех же условиях охлаждения сильно сокращаются. Когда наборы хромосом этих двух видов оказываются в одной гибридной соматической клетке и подвергаются охлаждению, то хромосомы S. tuberosum остаются палочковидными, a S. punae выглядят круглыми.
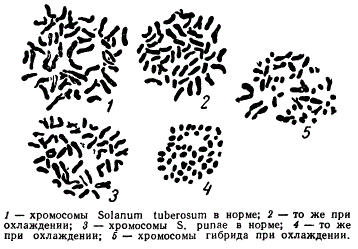
Поведение хромосом исходных видов в клетках гибрида Solanum tuberosum и S. punae
Следовательно, хромосомы разных видов сохраняют свои физиологические особенности даже в том случае, когда они находятся в другой цитоплазме.
Проблема совместимости и несовместимости геномов и цитоплазмы остаётся еще нерешенной, хотя и очень важной для понимания взаимосвязи ядра и цитоплазмы, а также роли аллополиплоидии в эволюции растений и животных.