Одним из явлений, видоизменяющих классическое расщепление по фенотипу 9 : 3 : 3 : 1 при дигибридном скрещивании, является неполное доминирование.
На примере моногибридного скрещивания у ночной красавицы (Mirabilis jalapa) и других объектов было показано, что при неполном доминировании одной из аллелей гена гибрид первого поколения оказывается промежуточным по проявлению альтернативных признаков. В потомстве такого гибрида в F2 происходит расщепление по фенотипу в отношении 1:2:1, что соответствует расщеплению по генотипу. Тот факт, что в данном случае гетерозиготное потомство отличается по внешнему виду от гомозиготного, дает возможность заключить, что доминирование есть результат взаимодействия пары аллелей, определяющих развитие данного признака. Иначе говоря, доминирование отражает преобладание одной аллели гена над другой (например, А над а, В над b и т. д.). Очевидно, что такое взаимодействие членов аллельной пары не всегда имеет своим результатом полное преобладание, т. е. полное доминирование.
Строго говоря, вероятно, генов с полным доминированием вообще не существует, так как ген,-как мы убедимся в дальнейшем, определяет не один какой-нибудь признак или свойство, а влияет на целый ряд признаков. Поэтому, говоря о доминировании или рецессивности какого-нибудь гена, мы условно прослеживаем лишь одну из сторон его действия — развитие одного признака.
В дигибридном скрещивании при неполном доминировании двух пар аллелей расщепление по фенотипу в F2 будет соответствовать формуле расщепления (1:2:1)2, или 1:2:2:4:1:2:1:2:1. Таким образом, мы будем иметь в F2 9 фенотипических классов, что соответствует числу генотипических классов — 32. Приведем два примера дигибридного скрещивания при неполном доминировании по двум парам аллелей у животного (курица) и растения (земляника).
У кур известен ген, вызывающий сильную курчавость оперения. Аллель Ã этого гена неполно доминирует над своей нормальной аллелью а, определяющей нормальное строение пера. Имеется и другая аллельная пара, находящаяся в другой паре гомологичных хромосом, которая вызывает так называемую разбрызганную окраску оперения — черные пятнышки на белом фоне. Эта окраска контролируется не полностью доминантной аллелью Ḃ. Рецессивная аллель b — определяет черную окраску. При скрещивании курицы с сильной курчавостью и черным оперением ÃÃbb с петухом, имеющим разбрызганную окраску и нормальное оперение ааḂḂ, гибриды первого поколения ÃаḂb оказываются промежуточными по выраженности этих признаков: у них проявляется слабая курчавость и голубая окраска. Во втором поколении такого дигибридного скрещивания расщепление по фенотипу наблюдается не в отношении 9:3:3:1, а в отношении 1:2:2:4:1:2:1:2:1.
Другой пример можно привести из опытов Т. С. Фадеевой, проведенных по анализу наследования признаков окраски ягоды и формы чашечки у земляники (Fragaria vesca). Красная окраска ягоды определяется гомозиготным состоянием аллели Ã, белая — аллели а. Другая аллельная пара в гомозиготном состоянии определяет нормальное строение чашечки ḂḂ, а рецессивное гомозиготное состояние гена bb приводит к образованию листовидной чашечки.
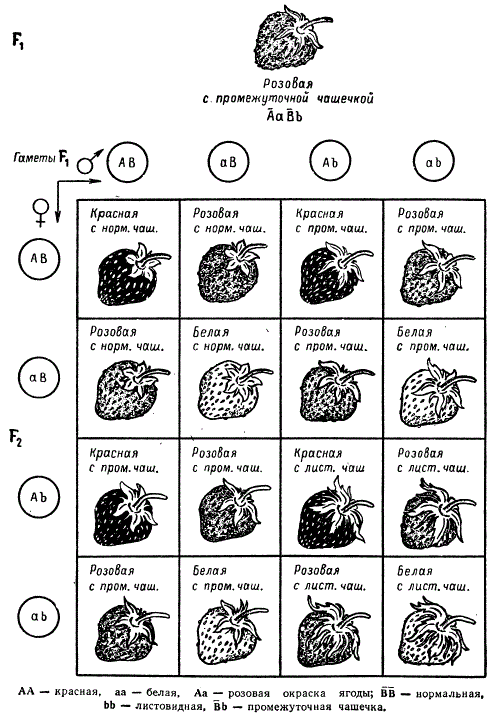
Наследование окраски ягоды и формы чашечки у Fragaria vesca при неполном доминировании в двух парах признаков
Гибридные растения первого поколения ÃаḂb имеют розовые ягоды и промежуточное строение чашечки. От самоопыления таких форм во втором поколении развиваются растения, которые составляют 9 различных фенотипических классов, соответствующих 9 классам генотипов. Статистически подтверждается совпадение теоретически ожидаемого расщепления по фенотипу с наблюдаемым в опыте: χ2 = 11,37, 0,20 > Р > 0,10.
Чаще встречаются такие случаи, когда в дигибридном скрещивании имеет место неполное доминирование лишь в одной из двух пар аллелей. Классическим примером в этом отношении является расщепление в F2 по признакам окраски и формы цветка у львиного зева (Antirrhinum majus). Красная окраска цветка у львиного зева определяется доминантной аллелью с неполным доминированием ее над рецессивной аллелью, обусловливающей белую окраску. Другая пара признаков касается формы цветка: нормальная — доминантная и пилорическая — рецессивная.
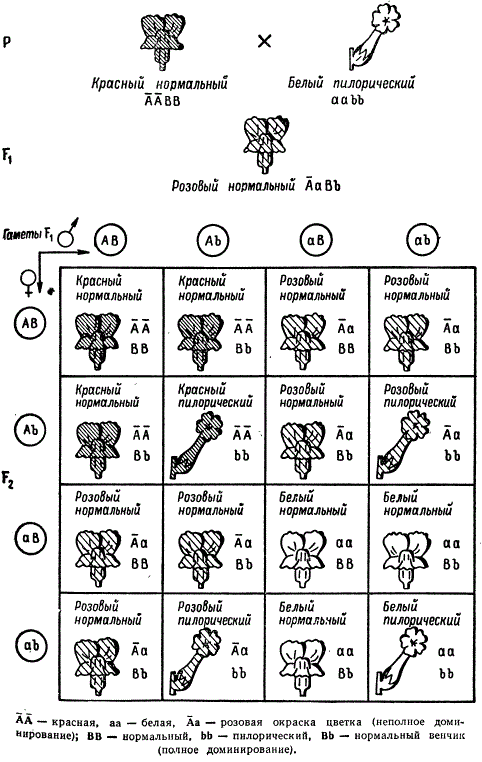
Наследование окраски и формы венчика у Antirrhinum majus при неполном доминировании в одной паре признаков
В данном дигибридном скрещивании расщепление в F2 дает 6 фенотипических классов: 3:6:1:2:3:1, причем красные цветки будут только в случае гомозиготного состояния доминантной аллели гена окраски, розовые — гетерозиготного ее состояния, а белые — гомозиготного состояния рецессивной аллели этого гена. По форме цветка промежуточных форм не будет, так как гетерозиготные и гомозиготные доминантные формы здесь не отличаются друг от друга.
Итак, неполное доминирование по одной или обеим парам аллелей генов в дигибридном скрещивании видоизменяет расщепление в по фенотипу. При полном доминировании в F2 будет 4 фенотипических класса, при неполном доминировании в одной из аллельных пар генов — 6 и при неполном доминировании в обеих аллельных парах — 9. В последнем случае расщепление по фенотипу полностью совпадает с расщеплением по генотипу.
Следует еще раз повторить положение, высказанное ранее, что неполное доминирование хотя и усложняет картину менделевского расщепления в дигибридном скрещивании, но ни в какой мере не противоречит законам Менделя, а подтверждает их. Явление неполного доминирования раскрывает новое содержание законов наследования. Если согласиться с тем, что признак обусловлен геном и характер расщепления в F2 определяется поведением хромосом в мейозе, то особенности расщепления при неполном доминировании по-новому раскрывают действие гена и характер его проявления. Характер проявления гена также может видоизменять расщепление по фенотипу.
Итак, явление независимого расщепления при полигибридном скрещивании может осуществляться при следующих условиях:
- нахождение учитываемых генов в негомологичных хромосомах; число их при этом не может превышать гаплоидное число хромосом у данного вида;
- равновероятное образование гамет всех сортов на основе случайного расхождения хромосом в мейозе;
- равновероятное созревание гамет всех типов;
- равновероятная встреча гамет при оплодотворении;
- равновероятная выживаемость зигот и взрослых организмов;
- относительная стабильность развития изучаемых признаков. Но все эти условия складываются под влиянием различных, постоянно и временно действующих факторов. В силу этого они могут изменяться, изменяя проявление закономерностей наследования.