До сих пор мы рассматривали физическую сторону влияния ионизирующей радиации на возникновение мутаций.
Но уже давно Г. Мёллером было установлено, что при облучении одной и той же дозой рентгеновых лучей зрелых и незрелых половых клеток частота выявляемых мутаций оказывается различной. Если одновременно облучить сперматозоиды и сперматогонии дрозофилы дозой 2000 р, то частота летальных мутаций в первом случае может быть около 2,5%, а во втором — около 0,5% или еще ниже.
Частота возникновения мутаций находится в линейной зависимости от дозы облучения. Но это действительно только для летальных мутаций в сперматозоидах и не реально для других стадий развития половых клеток.
Зависимость от дозы при облучении незрелых половых клеток оказывается сложной.
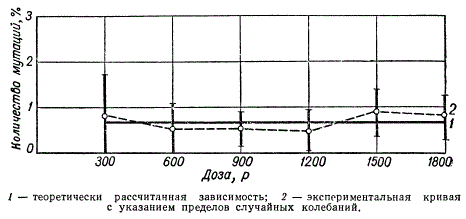
Частота рецессивных сцепленных с полом летальных мутаций у Drosophila при облучении незрелых половых клеток (сперматогониев) в зависимости от дозы рентгеновых лучей
При облучении сперматогоний дрозофилы относительно малыми дозами (100—300 р) частота летальных мутаций прямо пропорциональна дозе. То же явление наблюдается в другом диапазоне доз при облучении личинок (1500—3000 р). В диапазоне доз 600—1200 р частота мутаций от дозы не зависит, а зависит от биотических факторов. Такой характер кривой зависимости частоты мутаций в незрелых клетках обусловлен наличием двух типов сперматогониев: первичных А — нечувствительных и вторичных В — более чувствительных к радиации.
Для объяснения разной мутабильности зрелых и незрелых половых клеток Г. Мёллером уже давно были выдвинуты две альтернативные гипотезы. Согласно одной из них, высокая частота мутаций в сперматозоидах и пониженная — в сперматогониях могут быть следствием более высокой чувствительности сперматозоидов к ионизирующим излучениям. Согласно другой, эти стадии имеют одинаковую чувствительность, но незрелые клетки с возникшими в Них мутациями чаще подвергаются элиминации. Таким образом, последняя точка зрения допускает отбор среди зачатковых клеток. В 1938 г. М. Е. Лобашев высказал третье предположение, что незрелые половые клетки более чувствительны к ионизирующей радиации, чем зрелые, в силу чего элиминация среди первых выше, чем среди вторых.
Ни одна из этих гипотез до сих пор окончательно не отвергнута, но и не получила прямых доказательств. Возможно, что установленные различия мутабильности объясняются не первичными процессами, развивающимися при действии ионизирующих излучений, а специфичностью стадий клеток и вторичными процессами, разыгрывающимися в них после облучения. К последним можно отнести:
1) различную способность разорванных хромосом к восстановлению в незрелых (диплоидных) и зрелых (гаплоидных) половых клетках; предположительно эта способность выше в незрелых и диплоидных клетках;
2) различную степень элиминации сперматогониев и сперматозоидов с точковыми мутациями и хромосомными перестройками.
Главным барьером, отметающим хромосомные перестройки, образовавшиеся в сперматогониях, является мейоз, в профазе которого крупные хромосомные перестройки нарушают нормальную конъюгацию хромосом; мейоз «пропускает» лишь определенную группу мутаций — точковые и сравнительно мелкие хромосомные перестройки.
Это положение можно иллюстрировать следующими опытами. С помощью облучения рентгеновыми лучами у дрозофилы были получены летальные сцепленные с полом мутации. Затем эти мутации исследовались цитологически на гигантских хромосомах. Оказалось, что летальные мутации, возникшие в сперматозоидах и сперматидах в большом количестве (20—38%), связаны с крупными хромосомными перестройками — транслокациями, делециями и инверсиями; мутации же, учтенные в сперматогониях при тех же условиях облучения, ни в одном случае не были связаны с крупными хромосомными перестройками. Такого рода опыты убеждают в том, что мейоз играет роль своеобразного ОТК — отдела технического контроля в процессе гаметогенеза, отметающего клетки с крупными хромосомными перестройками.
Современные данные указывают на то, что наиболее высокой чувствительностью в отношении индуцирования мутаций ионизирующими излучениями обладают сперматиды, затем сперматозоиды, сперматоциты и менее всего сперматогонии. Причины такой градации чувствительности не выяснены.
В общей форме можно сказать, что ионизирующая радиация на разных стадиях сперматогенеза и оогенеза вызывает различный мутационный эффект. Это различие может быть обусловлено не только специфичностью действия агента и чувствительностью к нему клеток, но также особенностями состояния (спирализации) хромосом в разных фазах мейотического цикла и гаметогенеза у разных генотипов. Степень спирализации может обусловить разную способность воссоединения разорванных концов хромосом. Существенную роль могут играть и различия в интенсивности процессов элиминации мутировавших клеток, происходящих после облучения.