Цитоплазма живой клетки под световым микроскопом имеет вид прозрачной слизистой полужидкой однородной массы, не обладающей никакой внутренней структурой.
В нее погружены остальные органоиды клетки. Химический состав и физические свойства цитоплазмы очень сложны. Она не является однородным химическим веществом, а представляет собой организованную и постоянно меняющуюся систему из смеси разнообразных органических соединений, которые находятся частью в коллоидном состоянии, а частью в состоянии истинного раствора. Разнообразные минеральные соли, сахара и другие воднорастворимые соединения находятся в цитоплазме в истинном растворе. Белки, нуклеиновые кислоты, липоиды (жироподобные вещества), не растворимые в воде, образуют коллоидные растворы. Коллоидное состояние важнейших органических веществ цитоплазмы резко увеличивает поверхность соприкосновения компонентов при химических реакциях, протекающих с участием ферментов, и дает возможность (при наличии мембран) осуществлять в одно и то же время различного типа реакции в отдельных участках цитоплазмы.
Таким образом, по физическим свойствам цитоплазма представляет собой многофазный коллоидный раствор. Его существование связано с большим количеством воды — дисперсионной среды коллоида. Содержание воды в деятельной цитоплазме колеблется от 60 до 90%; в цитоплазме покоящихся семян и спор воды значительно меньше (5—15%). Большое количество воды объясняется главным образом тем, что в цитоплазме постоянно происходят сложнейшие химические реакции, для осуществления которых необходимо, чтобы реагирующие соединения находились в растворе.
Предполагают, что вода может находиться в связанном состоянии с другими веществами цитоплазмы и прежде всего с белками.
Веществом, определяющим характерные свойства и строение живого, являются белки — полимерные соединения, молекулы которых образуют в цитоплазме очень изменчивые мозаичные цепочки. Их можно разделить на две группы — простые белки (протеины) и сложные белки (протеиды). В состав протеидов входят собственно белок и небелковая группа, например, нуклеиновые кислоты, глюкоза, липоиды (жироподобные вещества). Наиболее важными из протеидов являются нуклеопротеиды, небелковая часть которых, так же как и белок, образована высокополимерными соединениями — нуклеиновыми кислотами. В цитоплазме обычно встречается только один тип нуклеиновых кислот—рибонуклеиновая кислота (РНК). Соединение белка с глюкозой дает глюкопротеиды, с липоидами — липопротеиды.
Цитоплазма ведет себя как коллоидная система, обратимо переходящая из золя в гель. Обычно она представляет собой гидрозоль, т. е. коллоидную систему с преобладанием дисперсионной среды — воды. При отдаче воды она может переходить в состояние геля, тогда начинает преобладать дисперсная фаза. Например, в покоящихся семенах цитоплазма находится в состоянии геля. При прорастании семян гидрофильные коллоиды сильно поглощают воду, набухают и цитоплазма переходит в состояние гидрозоля. Отдельные участки цитоплазмы даже одной клетки могут находиться в разном физическом состоянии. Более плотные мембраны цитоплазмы представляют собой гель, тогда как остальная часть находится обычно в состоянии золя?
При воздействии различных факторов («раздражителей») механического, химического или иного порядка цитоплазма легко меняет обычное для нормальных условий состояние гидрозоля и коагулирует (свертывается), при этом дисперсная фаза (белок и другие вещества) выпадает в виде хлопьевидного осадка. У большинства растений, находящихся в состоянии активной жизнедеятельности, цитоплазма необратимо коагулирует (погибает) от действия температур лишь немного выше 60° С. Однако в состоянии геля (покоящиеся семена) цитоплазма без губительных последствий может выдержать кратковременное действие температур даже ниже —100° С. Наряду с крайними температурами на цитоплазму разрушительно действуют также различные электролиты при высокой концентрации, электрический ток определенного напряжения й т. д. Явление коагуляции цитоплазмы наблюдается и при фиксации (быстром умерщвлении) растительных объектов, в результате которой цитоплазма становится видимой, так как в ней появляется зернистость. Одним из основных свойств цитоплазмы живой клетки является ее способность к движению. Движение цитоплазмы заметно главным образом во взрослых клетках, где она имеет вид постенного слоя, окружающего вакуолю. В этих клетках цитоплазма движется в одном направлении вокруг вакуоли, увлекая пластиды и митохондрии. Такое вращательное движение в соседних клетках происходит обычно в противоположных направлениях. Для клеток с тяжами цитоплазмы, пересекающими центральную вакуолю, характерно струйчатое движение. О механизме и функциях движения цитоплазмы до сих пор известно очень мало. Наиболее вероятно, что оно помогает передвижению необходимых веществ внутри протопласта.
Некоторые ученые считают, что движение цитоплазмы нормально не происходит и наблюдается только при повреждении клетки. Однако против этого говорят наблюдения, которые показывают, что температура, свет, электрический ток, различные вещества могут изменять скорость движения. Например, при действии электрического тока движение обычно останавливается.
Цитоплазма как живая система обладает свойством так называемой избирательной проницаемости: одни вещества, и прежде всего вода, легко проникают в нее, другие, в том числе растворенные в воде, цитоплазмой задерживаются.
Это находит свое выражение в явлении плазмолиза. Если погрузить живую растительную клетку в гипертонический раствор сахара или какой-нибудь соли, то эти вещества будут оттягивать воду из клетки, сами оставаясь вследствие избирательной проницаемости цитоплазмы вне клетки. Это оттягивание воды вызовет отставание эластичного протопласта от оболочки и съеживание его, что и называется плазмолизом.
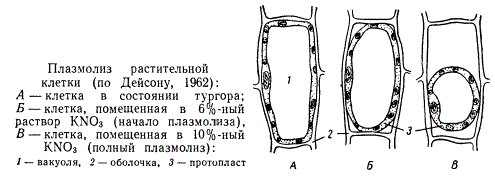
Плазмолиз растительной клетки
Плазмолиз обычно обратим, и при доступе воды или переносе клетки в гипотонический раствор вода снова энергично поглощается клеткой, и протопласт опять прижимается к оболочке. В результате клетка становится упругой, что обусловливает напряженное состояние тканей, а следовательно, и органов всего растения. Такое напряженное состояние клеток называется тургором.
Плазмолиз может быть показателем того, что данная клетка является живой, ибо отмершая клетка не плазмолизируется. Поэтому явление плазмолиза используется в анатомических исследованиях для суждения о том, является ли данная клетка живой или мертвой. Потеря тургора при плазмолизе вызывает явление завядания растения. При завядании на воздухе тонкостенные оболочки клеток сморщиваются и делаются складчатыми одновременно с протопластом.
В явлении избирательной проницаемости исключительная роль принадлежит двум пограничным слоям цитоплазмы: наружному слою, прилегающему к клеточной оболочке, называемому плазмалеммой (эктопластом), и внутреннему слою, отграничивающему внутреннюю поверхность цитоплазмы от вакуоли, называемому тонопластом. Доказательством особого состояния пограничных слоев, являющихся как бы барьером на пути веществ, служил опыт, при котором особым способом внутрь цитоплазмы впрыскивались различные органические красители. При этом красители не вымывались из цитоплазмы, а сохранялись в ней благодаря избирательно проницаемым пограничным слоем. Однако структура и химический состав плазмалеммы и тонопласта до недавнего времени оставались гипотетическими. Лишь электронный микроскоп позволил выяснить их строение. Оказалось, что как плазмалемма, так и тонопласт являются липопротеиновыми мембранами. Эти мембраны часто бывают асимметричными, что обусловлено различиями во взаимном расположении и строении молекул белков и липоидов, слагающих мембраны. Благодаря асимметрии одни вещества легче проходят в одном направлении, а другие — в противоположном. Поэтому на поверхности мембран обычно наблюдается неравномерное распределение ионов, создается разность электрических потенциалов, являющаяся, как предполагают, движущей силой многих биологических процессов. Плазмалемма и тонопласт функционируют как барьеры, ограничивающие свободную диффузию ионов, и контролируют состав и скорость проникновения молекул. Поэтому, например, корневой волосок, представляющий собой одну клетку, может содержать большое количество ионов калия и в то же время сохранять способность поглощать эти ионы из почвенного раствора, где концентрация их значительно ниже.
Однако не следует думать, что плазмалемма и тонопласт являются «глухими» барьерами, во всех своих точках оказывающими влияние на поглощение веществ. Были получены физиологические доказательства того, что в некоторых участках мембран вещества проходят свободно согласно законам диффузии. Эти участки настолько узки (5—10 Å), что их обычно не видно даже в лучший электронный микроскоп. Расположение и число этих участков не остаются постоянными.
Плазмалемма часто имеет волнистые контуры, иногда она образует складки, глубоко вдающиеся в цитоплазму и напоминающие узкие (менее 1 мк) канальцы. Эти канальцы обычно заполнены веществом клеточной оболочки, которое выделяется протопластом через плазмалемму. Иногда они являются электронно-пустыми, не заполненными веществом клеточной оболочки.
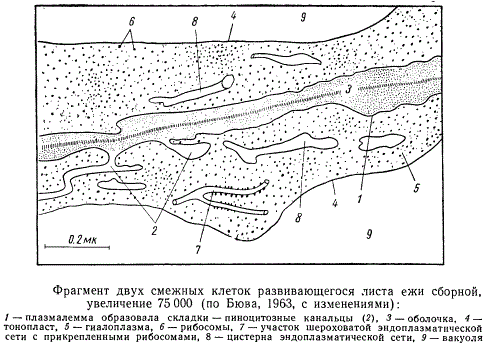
Фрагмент двух смежных клеток развивающегося листа ежи сборной
Возможно, такого типа канальцы служат для непосредственного поглощения клеткой растворов, в состав которых могут входить высокомолекулярные соединения — белки. Этот процесс, более изученный у животных клеток, получил название пиноцитоза. В процессе пиноцитоза раствор, оказавшийся внутри канальца, изолируется от внешней среды вследствие того, что концы мембран снаружи растут навстречу друг другу и затем сливаются в устье этого канальца. Раствор, находящийся в канальце, оказывается со всех сторон окруженным мембраной. Затем эта мембрана распадается, и вещество, заключенное в канальце, оказывается в цитоплазме. Хотя такой способ поглощения веществ у растительных клеток почти не исследован, но вполне возможно, что он существует.
Электронный микроскоп позволил обнаружить мембраны не только на поверхности, но и внутри цитоплазмы, которая на основании исследований в световом микроскопе считалась совершенно гомогенной. Эти мембраны отграничивают в цитоплазме разветвленную систему пузырьков, трубочек, уплощенных полостей («цистерн»), связанных между собой в более или менее непрерывную сеть, получившую название эндоплазматической.
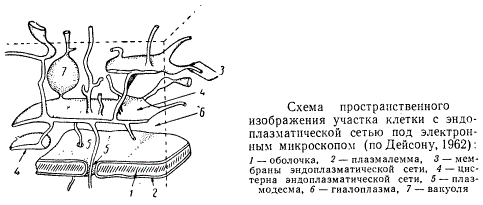
Схема пространственного изображения участка клетки с эндоплазматической сетью
Мембраны эндоплазматической сети разделяют цитоплазму на две обособленные фазы: первая представляет собой материал, находящийся внутри канальцев эндоплазматической сети, а вторая, располагающаяся снаружи от мембран, образует так называемую гиалоплазму (или матрикс) — основное вещество цитоплазмы. Содержимое канальцев и цистерн эндоплазматической сети в электронном микроскопе выглядит обычно гомогенным и прозрачным, реже в нем наблюдаются пучки фибриллярных элементов. По мнению некоторых ученых, по сети этих канальцев происходит движение различных веществ от плазмалеммы до мембраны ядра и обратно. Предполагают, что по этим канальцам белок может из цитоплазмы переходить в вакуоли и накапливаться там. В результате об эндоплазматической сети возникает представление как об особого рода циркуляционном аппарате цитоплазмы, направляющем и облегчающем обмен веществ между внешней средой и клеткой. Эндоплазматическая сеть — структура очень подвижная и непостоянная, ее мембраны непрерывно изменяются. Несмотря на их подвижность, мембраны эндоплазматической сети, как и все мембраны, вероятно, не возникают непосредственно из гиалоплазмы, а являются непрерывными во времени. Эндоплазматические мембраны могут быть связаны с ядерной мембраной и с плазмалеммой, но связь эта ввиду лабильности цитоплазмы постоянно меняется.
Гиалоплазма, находящаяся вокруг элементов эндоплазматической сети, представляет собой непрерывную недифференцированную фазу цитоплазмы, окружающую другие органоиды протопласта. Структура гиалоплазмы изучена очень слабо. В электронном микроскопе она имеет вид вещества неплотного, но обычно оно плотнее, чем содержимое элементов эндоплазматической сети. Иногда в нем можно наблюдать сеть из расположенных в беспорядке фибрилл диаметром около 200 Å. Об этих фибриллах тоже пока мало известно.

Участок гиалоплазмы из клетки флоэмной паренхимы тыквы
Иногда в периферической части гиалоплазмы, под плазмалеммой, в электронном микроскопе выявляются структуры, получившие название микротрубочек. Микротрубочки встречаются чаще всего в молодых, энергично растущих клетках (кончики корешков лука, тимофеевки, можжевельника, развивающиеся элементы древесины и луба) и располагаются перпендикулярно к их боковым стенкам и параллельно друг другу. Они обычно прямые или слегка скручены, имеют длину несколько микронов
и состоят из электронноплотной оболочки толщиной 55—70 Å и внутреннего светлого пространства диаметром 50—140 Å. Оболочка микротрубочек построена, в свою очередь, из 11—13 субъединиц — мельчайших структур, округлых на поперечном срезе и плотно прижатых друг к другу. Строение микротрубочек очень похоже на строение жгутиков одноклеточных организмов, служащих для их передвижения. Функция этих трубочек пока еще выяснена очень мало. Предполагают, что они играют роль в синтезе и ориентации целлюлозных микрофибрилл клеточной оболочки, а также в движении цитоплазмы и других органоидов клетки.
Довольно часто в гиалоплазме обнаруживаются участки с мельчайшей зернистостью. Эта зернистость вызывается скоплением гранул, имеющих округлую форму, диаметром около 150 Å, которые получили название рибосом (рибонуклеопротеидных гранул). Они богаты РНК и белком и на снимках получаются более темными, чем гиалоплазма, так как сильно рассеивают электроны. Функция рибосом была выяснена совсем недавно. Оказалось, что в них из аминокислот при участии РНК происходит синтез белковых молекул цитоплазмы. Рибосомы обычно не только свободно лежат в гиалоплазме,
но и в большом количестве прикрепляются к наружным поверхностям мембран эндоплазматической сети, рыхло покрывая их. Эндоплазматическая сеть с прикрепленными к ней снаружи рибосомами получила название «шероховатой», или гранулярной, а эндоплазматическая сеть без рибосом — «гладкой», или агранулярной. Рибосомы прикрепляются также к наружной поверхности ядерной мембраны, тогда как мембраны митохондрий, а также плазмалемма и тонопласт остаются гладкими. Помимо цитоплазмы рибосомы обнаружены внутри ядра, пластид и, возможно, митохондрий.
У молодых клеток гиалоплазма довольно плотная, очень богатая свободными рибосомами, а эндоплазматическая сеть не обильна и имеет вид немногочисленных разобщенных пузырьков. В клетках ‘более взрослых (дифференцированных) плотность гиалоплазмы уменьшается за счет гидратации, канальцы эндоплазматической сети удлиняются, расширяются и образуют многочисленные трубочки и цистерны, связанные друг с другом в единую сеть, пронизывающую всю цитоплазму. Мембраны эндоплазматической сети таких клеток обычно шероховатые, число свободных рибосом уменьшается. У клеток, находящихся в состоянии активного роста и синтеза клеточной оболочки, элементы эндоплазматической сети располагаются параллельно стенкам оболочки. В клетках старых и малоактивных гиалоплазма еще более гидратирована, объем эндоплазм этической сети вновь уменьшается, мембраны становятся гладкими (без рибосом), а свободные рибосомы встречаются очень редко.