Под явлением наследственности мы понимаем широкий биологический процесс материальной и национальной преемственности между поколениями.
Основу этой преемственности составляет механизм клеточного деления как в случае половых, так и соматических клеток. Очевидно, что любые структуры клетки, которые воспроизводятся и распределяются при делении в дочерние клетки, могут передавать наследственную информацию, следовательно, задача исследователей и заключается, чтобы изучить значение всех структурных элементов клетки общей системе ее деятельности и воспроизведения.
Уже давно генетики встречались с многочисленными фактами, которые не укладывались в рамки хромосомной теории наследственности. Издавна известно, что различие гибридов от реципрокных скрещиваний при отдаленной гибридизации дает основание говорить неравном участии женских и мужских половых клеток в образовании гибридного организма, и наиболее правдоподобно эти различия отнести за счет неравного количества цитоплазмы в яйцеклетке и сперматозоиде.
Признаки, за наследование которых ответственны элементы цитоплазмы, должны передаваться главным образом по материнской линии. Поэтому для установления факта наследовании какого-либо признака через цитоплазму необходимо выявление различий в реципрокных скрещиваниях. Это первый этап. Следующим этапом анализа цитоплазматической наследственности являются возвратят скрещивания гибрида с отцовской формой для замещения всех материнских хромосом отцовскими. Если и при этом сохранится передача признака по материнской линии, цитоплазматический характер наследования его можно считать доказанным.
Под явлением цитоплазматической наследственности (ЦН) следует понимать наследование признаков и свойств организма, детерминированных элементами цитоплазмы и ее органоидами.
Основоположниками изучения цитоплазматической наследственности являются немецкие генетики К. Корренс и Э. Бауэр.
Прежде чем перейти к анализу фактов собственно цитоплазматической наследственности, необходимо, во-первых, установить соотношение роли ядра и цитоплазмы в наследственности и, во-вторых, рассмотреть ряд явлений, которые имитируют цитоплазматическую наследственность по своему проявлению, но не относятся к таковой: это случаи наследования через различные инфекции цитоплазмы, явления длительных модификаций и предетерминации цитоплазмы и др.
Для того чтобы взвесить значение отдельных элементов цитоплазмы в наследственности при половом размножении, необходимо, во-первых, определить те свойства, которыми они должны обладать, чтобы осуществлять функцию передачи информации от одного клеточного поколения к другому, во-вторых, определить те структуры цитоплазмы, которые обладают этими свойствами.
Для того чтобы цитоплазма и ее структурные элементы обладали свойством передачи информации в поколениях, они должны:
- относиться к составным, жизненно необходимым для клетки структурам,
- обладать способностью к репродукции,
- распределяться при клеточном делении,
- иметь способность изменяться и устойчиво передавать эти изменения в поколениях.
Роль органоидов цитоплазмы
В цитоплазме клетки имеются различные органоиды: центриоли, пластиды, митохондрии и рибосомы.
Трудностями в изучении генетического значения структурных элементов цитоплазмы являются, во-первых, сложность установления морфологической индивидуальности их в клетке, во-вторых, отсутствие для них каких-либо маркеров, подобных генам в хромосомах. Роль отдельных органоидов цитоплазмы в наследовании зависит от характера их дискретности и способности свободно комбинироваться и строго распределяться при клеточном делении.
Центриоль имеется в клетках животных и низших растений, у высших растений ее нет. Центриоль содержит ДНК, обладает свойством делиться в профазе митоза и мейоза, следовательно, ее структура способна репродуцироваться. Центриоль имеет прямое отношение к образованию аппарата веретена деления, но в передаче информации в ряду клеточных делений ее роль не ясна и характер ее изменчивости неизвестен.
Митохондрии состоят из белков, специфических РНК и ДНК и фосфолипидов. Кроме того, митохондрии содержат комплекс ферментов, способных расщеплять углеводы, жирные кислоты и аминокислоты до углекислоты и воды. Они чаще сосредоточиваются в местах с наиболее интенсивным обменом веществ в клетке. Предполагается, что в растительных клетках они связаны с образованием пластид. Изолированные из клеток путем гомогенизации митохондрии сохраняют способность расщеплять углеводы, жирные кислоты и аминокислоты. Механизм их образования неизвестен, но показано, что высокая их биохимическая активность тесно связана с действием генов. Исследования последних лет все больше убеждают в том, что митохондрии принимают участие в наследственной передаче некоторых функций клетки, в частности дыхания.
Рибосомы встречаются во всех клетках. Они ответственны за синтез белков первичной структуры.
Наиболее изученными в генетическом плане органоидами цитоплазмы являются пластиды, представляющие собой своеобразные Лаборатории синтеза углеводов растительного организма.
Для установления истинной роли структурных элементов цитоплазмы в передаче наследственной информации важно знать, воспроизводятся ли они при клеточном делении путем автономной репродукции так же, как хромосомы, или они возникают заново в процессе жизнедеятельности клетки под контролем ядра, т. е. генотипа. Однако до сих пор эти вопросы еще очень мало исследованы.
Пластидная наследственность
Пластиды состоят из специфических белков, ДНК и РНК.
В зависимости от содержащихся в них пигментов пластиды подразделяются на хлоропласты (зеленый пигмент), хризопласты (желтый и коричневый), (реопласты (бурый), родопласты (красный) и лейкопласты (бесцветные).
Количество пластид на клетку у разных видов высших и низших растений колеблется от 1 до 100 и более. Виды с одной пластидой в клетке встречаются у жгутиковых и мхов, с двумя пластидами — среди диатомовых водорослей.
Внутренняя структура пластид сложная — в них найдены мембранная система типа эндоплазматического ретикулума и рибосомы. В пластидах содержатся РНК и ДНК. ДНК пластид по составу оснований отличается от ДНК хромосом, а РНК пластидных рибосом — от рибосомальной РНК цитоплазмы.
Некоторые авторы считают, что генетическая информация пластид заключена в их ДНК.
Установлено, что пластиды размножаются делением и расходятся в дочерние клетки во время митоза. Найдены примеры (в роде Cylindrocystis), когда в ходе оплодотворения сливаются две клетки, каждая из которых несет по две пластиды, в результате чего образуется зигота с 4 пластидами. После мейоза каждая из гаплоидных клеток получает по одной пластиде, которая делится еще один раз. В результате образующаяся гамета содержит две пластиды. Данная картина представляет собой своеобразную редукцию числа пластид, приуроченную к процессу мейоза.
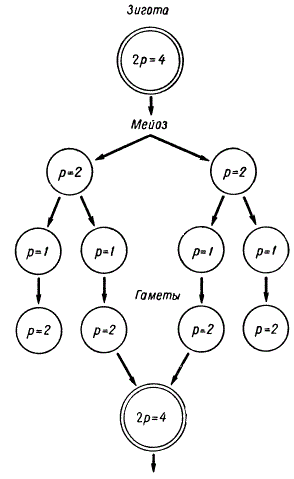
Схема редукции числа пластид у Cylindrocystis, связанной с мейозом и оплодотворением
Клетки, утратившие пластиды, не способны их образовывать заново. Так, одноклеточная Euglena rnesmily имеет около 70—100 хлоропластов, но если ее содержать в темноте, то репродукция хлоропластов затормозится, и по мере деления клеток могут возникнуть особи, совершенно лишенные пластид. Они не способны образовать заново пластиды; в культуре они поддерживаются на искусственной питательной среде.
Генетические свойства пластид были установлены уже давно. Совокупность пластид клетки как структур, способных передавать наследственную информацию, была названа О. Реннером пластидомом. Из всех структурных элементов цитоплазмы растений, с которыми можно связывать передачу некоторых свойств и признаков материнского организма потомству, пластиды наиболее удобны для анализа, так как в большинстве случаев они являются четко различимыми структурами, обладающими целым рядом морфологических особенностей. Они способны к скачкообразным изменениям — пластидным мутациям, которые в дальнейшем четко воспроизводятся.
О первых фактах пластидной наследственности сообщили Э. Баур и К. Корренс еще на заре развития генетики (в 1908 г.). Э. Баур изучал наследование белой пестролистности у герани Pelargonium zonale. Он установил, что пестролистные растения являются химерными по своему строению.
Химеры образуются трех типов: 1) сочетание разнокачественных тканей 2) наличие разнокачественных групп клеток в пределах одной и той же ткани и 3) сочетание различающихся органоидов внутри одной клетки и неравномерное распределение их при клеточном делении.
У герани известны пестролистные растения, которые имеют в эпидермальном и субэпидермальном слоях точек роста группы клеток с пластидами, не способными к образованию хлорофилла, тогда как в центральных слоях клетки додержат нормальные пластиды. Вследствие этого иногда на растении образуются чисто зеленые или совершенно белые ветви. Семена, порученные с белых ветвей, дают нежизнеспособные сеянцы.
При опылении цветков с пестролистных ветвей пыльцой от цветков с зеленых ветвей и при реципрокном скрещивании результаты получаются различными. В первом случае (пестролистное X зеленое) гибридные растения развиваются пестролистными, зелеными и белыми (гибнут). При реципрокном скрещивании (зеленое X пестролистное) в потомстве все растения оказываются зелеными. На основании этого был сделан вывод о том, что наследование пестролистности связано с передачей и распределением при клеточных делениях двух типов пластид — зеленых и неокрашенных. Данные скрещиваний показывают, что передача пластид осуществляется через яйцеклетки.
Развитие же белых или зеленых частей растений из зиготы, содержащей пластиды обоих типов, определяется скоростью и способом воспроизведения этих разных пластид и их распределением — отмешиванием в ходе клеточных делений.
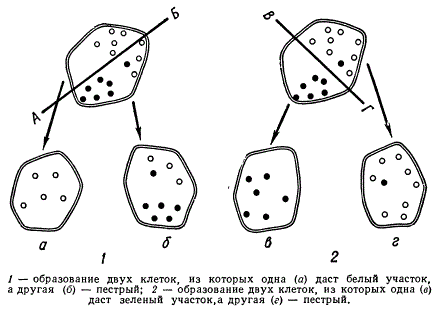
Схема случайного распределения белых и зеленых пластид при клеточном делении
Клетки, получившие при таком отмешивании только зеленые пластиды, дают далее зеленые участки тканей, а клетки, получившие только неокрашенные пластиды, — белые участки. Если при делении клетки клеточная оболочка пройдет по линии АВ, то образуются две клетки, которые дадут два участка (белый — а и пестрый — б), при разделении по линии ВГ — зеленые и пестрые (в, г). При этом граница между белыми и зелеными зонами тканей растения не резкая, ибо некоторые клетки содержат оба типа пластид. Такой же тип наследования пестролистности, как у Pelargonium zonale, известен и у ряда других растений: ржи, львиного зева, кукурузы, примулы и др.
К. Корренс описал другой тип наследования пестролистности. Объектом его работ была ночная красавица Mirabilis jalapa. Среди Mirabilis есть растения, которые могут иметь чисто зеленые, белые и пестролистные ветви, причем граница между зеленой и белой зонами в отличие от герани всегда резкая. Это объясняется тем» что в каждой клетке Mirabilis могут быть пластиды только одного типа — либо зеленые, либо неокрашенные. В силу этого отмешивание пластид при клеточном делении, которым можно объяснить пестролистность у герани, в данном случае не может иметь места. Корренс производил опыление цветков с белых, зеленых и пестролистных ветвей пыльцой от цветков всех трех типов.
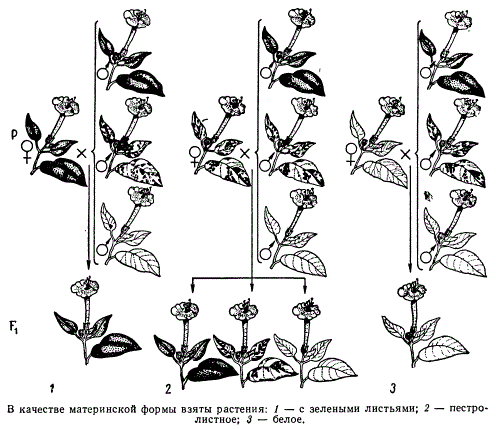
Наследование пестролистности типа status albomaculatus (белопятнистость) у Mirabilis jalapa
У гибридных растений он обнаружил три типа окраски листьев: белые, зеленые и пестрые. Анализ этих данных показал, что наследование пестролистности у этого растения осуществляется полностью по материнской линии.
Очевидно, у Mirabilis наличие зеленых или неокрашенных пластид в клетке обусловлено какими-то различными состояниями цитоплазмы. У чисто белых и полностью зеленых растений эти различные состояния цитоплазмы оказываются устойчивыми. В клетках пестролистных растений цитоплазма характеризуется лабильным состоянием. В ходе развития растения цитоплазма клеток может переходить в любое из двух устойчивых состояний, давая чисто белые или чисто зеленые ветви, или оставаться лабильной, давая Пестролистные ветви.
Пестролистность, передающаяся только яйцеклетками, но не пыльцой, была названа К. Корренсом status albomaculatus (белопятнистостью). Сходные явления обнаружены также у львиного зева, подорожника, хмеля и других растений.
У некоторых растений наблюдается закономерная изменчивость проявления пестролистности в ходе онтогенеза. Например, у пестролистных растений хмеля (Humulus japonica) семядоли всегда бывают чисто зелеными, а на развивающихся листьях побега пестролистность тем сильнее, чем позднее формируется лист. На боковых побегах проявление этого признака характеризуется такой же последовательностью.
Развитие пестролистности у подорожника в значительной степени зависит от температуры и, возможно, от светового режима. При низких температурах пестролистность развивается значительно сильнее, а повышение температуры приводит к ее ослаблению или даже полному исчезновению.
Изменчивость пестролистности в индивидуальном развитии позволяет произвести онтогенетический анализ развития признаков, наследование которых связано с плазмоном и пластидомом. Такой анализ представляет собой метод для установления закономерностей в распределении цитоплазматических структур, передающих наследственную информацию, и в способе их действия.
Установлено, что, как правило, свойства пластид определяются непосредственно ядерными генами и наследование пестролистности при этом осуществляется в соответствии с установленными закономерностями ядерной наследственности, но измененные пластиды передаются вместе с цитоплазмой.
Известны также факты, когда заболевание пластид вызывается различными вирусами. Передача пестролистности в последовательных поколениях при этом имитирует цитоплазматическую наследственность. Однако такие инфекции представляют совершенно особую группу явлений.
Цитоплазматическая мужская стерильность
Одним из самых ярких примеров цитоплазматической наследственности представляется явление цитоплазматической мужской стерильности (ЦМС), обнаруженное у многих растений, — у кукурузы, лука, свеклы, сорго, льна и др.
Цитоплазматическая мужская стерильность у кукурузы была открыта в тридцатых годах в СССР М. И. Хаджиновым и в США — М. Родсом.
Цитоплазматическая мужская стерильность наиболее полно изучена у кукурузы. Кукуруза является однодомным растением, женские цветки которого собраны в початок, мужские — в метелку. У некоторых сортов кукурузы были обнаружены растения, имевшие в метелках недоразвитые пыльники, часто совершенно пустые, а иногда с недоразвитой стерильной пыльцой.

Нормальная (слева) и стерильная (справа) метелки кукурузы
Как оказалось, этот признак определяется особенностями цитоплазмы. Опыление растений с мужской стерильностью нормальной пыльцой с других растений в большинстве случаев дает в потомстве растения со стерильной пыльцой. При повторении этого скрещивания в течение ряда поколении признак мужской стерильности сохраняется, передаваясь по материнской линии. Даже тогда, когда все 10 пар хромосом таких стерильных по пыльце растений замещаются хромосомами от растений с нормальной фертильной, пыльцой, признак мужской стерильности сохраняется. Все это послужило убедительным доказательством того, что наследование данного признака осуществляется через цитоплазму. Такое явление и было названо цитоплазматической мужской стерильностью. Цитоплазма, обусловливающая стерильность пыльцы, была обозначена символом цитS (стерильная цитоплазма), а цитоплазма растений с фертильной пыльцой — символом цитN (нормальная цитоплазма). В дальнейших исследованиях было обнаружено, что генотип растения может оказывать определенное влияние на действие стерильной цитоплазмы. Цитоплазма цитS может обусловить стерильность пыльцы только при наличии в генотипе растения рецессивных генов rf в гомозиготном состоянии цитS rfrf. Если же этот ген представлен доминантной аллелью Rf, то растение цитS RfRf или цитS Rfrf имеет нормальную пыльцу. Ген Rf является, таким образом, восстановителем фертильности пыльцы. Следовательно, фертильную пыльцу могут иметь и растения цитN rfrf и цитN RfRf или Rfrf, и цитS Rf Rf или Rfrf, а полностью стерильную только растения цитSrfrf. Многократное повторение скрещивания растений цитSrfrf х цитNrfrf дает потомство с полностью стерильной пыльцой. И только в случае скрещивания цитSrfrf х цитSRfRf (или цитNRfRf) может быть получено потомство, где все растения будут иметь нормальную пыльцу, несмотря на наличие цитоплазмы цитS.
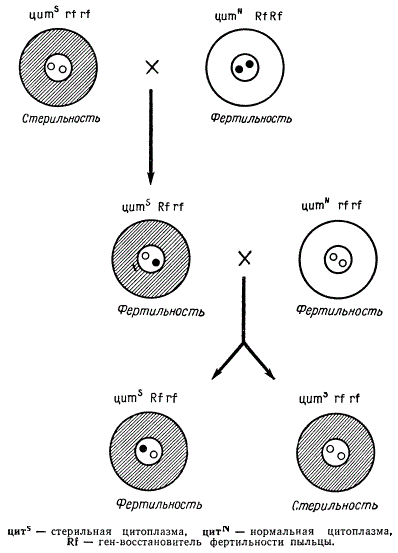
Схема наследования цитоплазматической мужской стерильности
Следует еще раз подчеркнуть, что ген Rf не изменяет структуру и специфичность цитоплазмы, а лишь тормозит проявление ее действия. В настоящее время обнаружен еще целый ряд генов, способных оказывать влияние на действие стерильной цитоплазмы.
Исследование цитоплазматической мужской стерильности представляет яркий пример успешного генетического анализа ядерно-цитоплазматических отношений.
Цитоплазматическое наследование дыхательной недостаточности
Ядро и цитоплазма имеют как одинаковые, так и различные ферменты, определяющие в них разные метаболические процессы.
Однако дыхательные ферменты обнаружены только в цитоплазме и они связаны с митохондриями. В изолированных ядрах клеток дыхание не установлено.
У некоторых грибов (пекарские дрожжи, нейроспора) были обнаружены дефектные изменения дыхательной системы (дыхательная недостаточность). Дыхательная недостаточность у грибов обусловлена необратимыми наследственными изменениями структуры и функции митохондрий. При этом изменении они утрачивают активность цитохромоксидазы. В высокоочищенных митохондриях дрожжей найдена собственная ДНК.
Б. С. Эфрусси обнаружил штаммы пекарских дрожжей (Saccharomyces cerevisiae), которые спонтанно образуют мелкие карликовые колонии (petites) с дыхательной, недостаточностью. Поскольку они возникают при вегетативном размножении гаплоидных дрожжей, они были названы «вегетативными карликовыми». Эти мутанты при скрещивании между собой не дают диплоидной зиготы и аскоспор.
Наряду с вегетативными карликовыми колониями была обнаружена мутантная форма, по фенотипу — росту и дыхательной недостаточности — сходная с первой. Но эта карликовая форма давала расщепление по указанному признаку — карликовости, такое, как будто данный фенотип определяется мутацией одного ядерного гена. Эта форма была названа расщепляющимся карликовым штампом.
Так как у дрожжей при скрещивании и образовании зиготы сливаются целые клетки — цитоплазмы и ядра, то в этом случае можно оценить раздельно роль цитоплазмы и ядра в наследовании при скрещивании данных мутантов с нормальной формой.
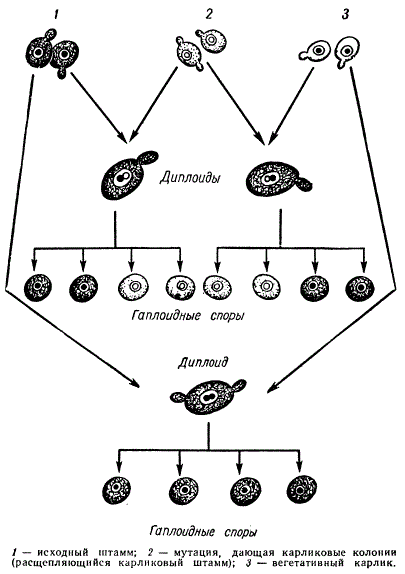
Схема генетического анализа вегетативного и расщепляющегося штаммов у Saccharomyces cerevisiae
Приведена схема генетического анализа вегетативного и расщепляющегося карликового штаммов. Из этой схемы видно, что фенотип расщепляющейся карликовости детерминирован ядерным геном, поскольку при скрещиваниях наблюдается расщепление аскоспор в отношении 1:1. При скрещивании вегетативных Карликов и нормальных дрожжей диплоидная зигота не дает расщепления — из аскоспор не появляются мелкие колонии. Следовательно, у нормальных дрожжей и вегетативных карликовых геномы одинаковы, различия лишь в цитоплазме. Расщепления по типу цитоплазмы в мейозе не происходит.
В данном случае, когда сливается цитоплазма обеих родительских клеток, а наследование осуществляется по материнской линии, факт цитоплазматической наследственности приобретает особую убедительность. Таким образом выяснилось, что вегетативные карликовые мутанты являются цитоплазматическими мутантами, возникновение которых не обусловлено каким-либо ядерным геном, хотя частота спонтанного возникновения таких мутантов зависит от генотипа материнской клетки (штамма).
В последнее время было открыто новое явление — цитоплазматическая трансформация (ЦТ) с помощью митохондрий пекарских дрожжей. Для доказательства цитоплазматической трансформации спирали клетки дрожжей с дыхательной недостаточностью. Желудочным соком улиток растворяли у них оболочки и получали, клетки без оболочки — сферопласты. Затем из дрожжей дикого типа получали очищенную фракцию митохондрий с нормальной дыхательной функцией и добавляли ее в суспензию сферопластов petites. После инкубации совместно с фракцией митохондрий из сферопластов развивались колонии дикого типа с частотой 2—2,5%.
Этот тип трансформации открывает широкие перспективы дли изучения взаимосвязи соматических клеток в ткани in situ и в культуре тканей. Природа трансформирующего фактора остается неясной — эффект цитоплазматической трансформации можно отнести за счет проникновения целых митохондрий или их фрагментов, то можно предполагать также, что эффект трансформации достигается здесь с помощью изолированной ДНК, как это было уже установлено для бактерий.
У нейроспоры также известен четкий факт наследования дыхательной недостаточности через цитоплазму. У нейроспоры есть штаммы, отличающиеся от нормальной формы дикого типа замедленным ростом и дефектами в дыхательной системе (нарушение цитохромной системы), — так называемые штаммы року.
Как известно, оплодотворение зрелых плодовых тел — протодеритециев осуществляется микроконидиями, которые почти лишены цитоплазмы. Диплоидной фазой в жизненном цикле нейроспоры является только зигота; она испытывает редукционное деление с последующим митозом и дает восемь аскоспор, развивающихся в сумке. При реципрокных скрещиваниях штамма року со штаммом дикого типа мицелии, развивающиеся из аскоспор — продуктов редукционного деления гибридной зиготы, всегда обнаруживают признаки той родительской формы, ко второй принадлежал протоперитеций. Это дает основание сделать вывод о том, что в данном случае наследственная передача осуществляется по материнской линии через цитоплазму. Такой характер наследования сохраняется как в последующих аналогичных скрещиваниях, так и в большом числе последовательных вегетативных поколений.