Определение цветка покрытосеменных растений — задача сложная и до сих пор окончательно не решенная.
Наиболее распространенное и общепринятое определение такое: цветок есть укороченный побег с ограниченным ростом, несущий спорофиллы — тычинки и плодолистики и в типичном случае стерильные цветолистья, составляющие околоцветник.
Так как гипотезы о происхождении цветка покрытосеменных растений, его эволюции с должной подробностью рассматриваются в соответствующих разделах морфологии и систематики растений, то, не останавливаясь на них, перейдем к рассмотрению анатомии различных органов цветка покрытосеменных.
Следуя «классической» гипотезе происхождения и строения цветка, чашелистики, лепестки, тычинки и пестики рассматривались как метаморфизированные листья и поэтому иногда назывались цветолистиками. Наиболее определенно признаки листовой природы обнаруживаются в листочках околоцветника: чашелистиках и лепестках.
Среди громадного разнообразия организации цветков достаточно отчетливо выделяются две основные категории цветков: актиноморфные (их также называют правильными) и зигоморфные (которые называют неправильными). Актиноморфными называются такие цветки, через которые можно провести несколько плоскостей симметрии. Зигоморфный цветок имеет только одну плоскость симметрии.
Каждый цветок состоит из более или менее четко выделяющихся циклов, или мутовок, причем обычно таких циклов пять: два цикла околоцветника, два цикла андроцея (тычинок) и один цикл гинецея, состоящий из плодолистиков. Каждый цикл состоит или из трех (у однодольных), или из четырех, пяти и более членов.
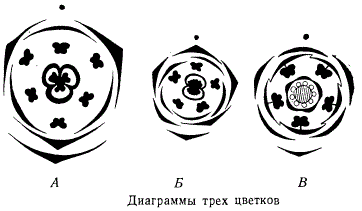
Изучая организацию цветков различных растений, как актиноморфных, так и зигоморфных, легко убедиться, что строение цветков подчиняется определенной закономерности. Чтобы более отчетливо представить себе тип каждого цветка, строят диаграммы, на которые наносят в определенной проекции все части цветка в их взаимном расположении. Выводятся также формулы цветка, передающие в цифрах общий состав каждой из его мутовок.
Для каждого цветка можно составить свою диаграмму, с той или иной точностью передающую характерные черты, если не вида цветка, то по крайней мере рода, к которому растение относится. Примеры диаграмм представлены на рисунке — диаграмма цветка однодольного типа лилейных, изображает организацию трехмерного актиноморфного пятициклового цветка, состоящего из круга наружного околоцветника, круга внутреннего околоцветника, двух кругов тычинок и одного круга, или цикла, трехчленного гинецея. Формула такого цветка: К3С3А3+3G3. Б — диаграмма пятимерного цветка двудольного из подкласса раздельнолепестных с двухцикловым околоцветником, одноцикловым андроцеем и двухгнездной завязью (гинецеем). Формула цветка: K5C5A5G2; В — диаграмма пятимерного цветка двудольного с двухцикловым околоцветником, причем внутренний цикл — спайнолепестный с приросшими к лепесткам тычиночными нитями и одногнездным гинецеем.
Морфологически чашелистики как члены наружного круга околоцветника нередко обнаруживают различно выраженные переходы через промежуточные формы от настоящих листьев. У одних растений чашелистики по внешней и внутренней организации напоминают верхушечные листья стебля, а у других совершенно неотличимы от лепестков, составляя с последними так называемый простой околоцветник вместо обычно формирующегося двойного околоцветника, в котором чашелистики и лепестки резко друг от друга отличаются. У одних растений чашелистики эфемерны, быстро опадают после цветения или даже до него. Но есть также немало растений, в цветках которых чашелистики не только остаются, но и принимают участие в формировании плода. Иногда чашелистики сильно разрастаются (например, у физалис), изменяют первоначальную зеленую окраску и т. д. Эпидермис чашелистиков обычно снабжен относительно тонкой кутикулой и устьицами, мезофилл, как правило, состоит из хлорофиллосной паренхимы. Но устьиц в эпидермисе чашелистиков по сравнению с числом устьиц на настоящих листьях мало, мезофилл обычно не дифференцируется на палисадную и губчатую паренхимы, целиком состоя из очень рыхло расположенных клеток с большими межклетниками, нередко развивающимися в обширные воздухоносные полости или ходы.
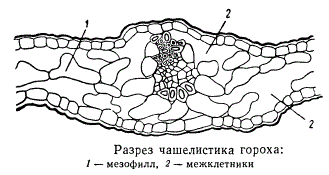
Сосудисто-волокнистая система чашелистиков редуцирована, без вторичных утолщений, со слабым развитием или полным отсутствием лубяных волокон. Часто отсутствует также колленхима. Вообще в своем развитии механическая система в чашелистиках значительно уступает механической системе листьев. У некоторых растений чашелистики после цветения одревесневают, становясь жесткими и даже превращаясь в колючкоподобные образования.
Еще большую степень изменений по сравнению с листьями проявляют лепестки. В отличие от чашелистиков лепестки значительно реже бывают зелеными, в особенности у травянистых растений. На лепестках тоже образуются устьица, хотя в весьма незначительных количествах и чаще всего не вполне развитые. Если мезофилл лепестков содержит хлоропласты, то такие же хлоропласты есть и в замыкающих клетках устьиц. При отсутствии хлоропластов в мезофилле их нет и в замыкающих клетках устьиц. Эпидермис лепестков покрыт кутикулой, у разных растений различной толщины, но в общем не достигающей значительной мощности, за некоторым исключением.
 Эпидермальные клетки лепестков весьма разнообразно смыкаются друг с другом, образуя всевозможные складки. При этом в различных местах одного и того же лепестка очертания эпидермальных клеток могут быть различными. Иногда же между эпидермальными клетками образуются межклетники, достигающие относительно значительных размеров и затянутые кутикулой.
Эпидермальные клетки лепестков весьма разнообразно смыкаются друг с другом, образуя всевозможные складки. При этом в различных местах одного и того же лепестка очертания эпидермальных клеток могут быть различными. Иногда же между эпидермальными клетками образуются межклетники, достигающие относительно значительных размеров и затянутые кутикулой.
В то время как на чашелистиках образуются настоящие волоски, простые или железистые, эпидермис лепестков преимущественно образует выросты, имеющие вид либо сосочков, либо несколько более удлиненных мешковидных образований, нередко содержащих различные пигменты. От присутствия таких сосочков поверхность лепестков приобретает бархатистость. Лепестки бывают весьма разнообразной консистенции: тонкие и нежные, как, например, у льна, черемухи и толстые мясистые с признаками некоторой суккулентности структуры, как у некоторых видов седума и у королевской лилии. В зеленых и толстых лепестках мезофилл напоминает ткань листьев сильнозатененных растений: палисадной паренхимы нет, клетки расположены рыхло, только форма клеток мезофилла толстых лепестков обычно округлая, а в листьях теневых растений мезофилл чаще состоит из клеток, образующих отростки различной величины и формы. В белых или окрашенных лепестках осуществляются различные формы редукции мезофилла, доходящие в лепестках некоторых растений до того, что весь мезофилл состоит из одного слоя клеток, утолщаясь лишь там, где проходят жилки.
Клетки мезофилла обычно тонкостенные, целлюлозные, но изредка в лепестках встречаются склереидные анатомические элементы, например у камелиевых. В лепестках встречаются и различные вместилища секретов и т. п. У актиноморфных цветков все лепестки построены одинаково, у зигоморфных структура одних лепестков может значительно отличаться от структуры других.
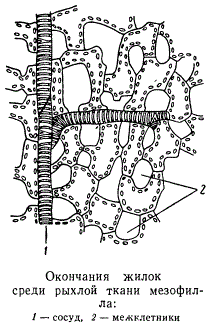 Проводящая система лепестков также значительно редуцирована по сравнению с проводящей системой листьев. Вторичные утолщения отсутствуют, сосуды примитивной структуры, лубяных волокон чаще всего нет. Обычно жилки лепестков состоят из очень небольшого количества спирально-кольчатых трахеид, свободно оканчивающихся среди мезофилла. Таких обкладок вокруг жилок, какие имеются в листьях, в лепестках часто не бывает.
Проводящая система лепестков также значительно редуцирована по сравнению с проводящей системой листьев. Вторичные утолщения отсутствуют, сосуды примитивной структуры, лубяных волокон чаще всего нет. Обычно жилки лепестков состоят из очень небольшого количества спирально-кольчатых трахеид, свободно оканчивающихся среди мезофилла. Таких обкладок вокруг жилок, какие имеются в листьях, в лепестках часто не бывает.
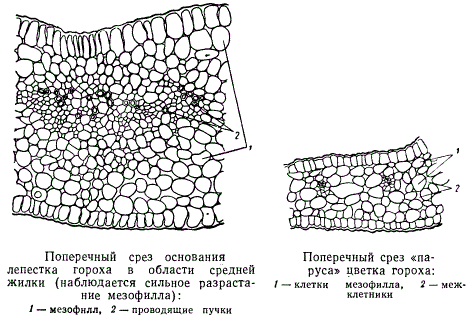
Пластинки лепестков, особенно тонких, редко бывают одинаковой толщины во всех местах. Наибольшая толщина лепестка у основания, в месте прикрепления его к цветоложу. Кроме того, по определенным местам лепестка встречаются весьма характерные более или менее сильно выраженные утолщения. Образуются они часто там, где проходит средняя жилка или по бокам ее, в базальной части лепестка.
Клетки паренхимы мезофилла лепестка часто бывают округлыми, но нередко образуют неправильные отростки. Контакт между клетками осуществляется через отростки. В результате в мезофилле лепестка создается хорошо развитая система межклетных ходов. Паренхима подобного строения называется копулирующей. В мясистых лепестках и в утолщенных частях тонких лепестков копулирующей паренхимы не образуется, там — клетки округлые.
Лепестки могут быть пигментированными (окрашенными) и неокрашенными. Неокрашенные лепестки имеют снежно-белый цвет. Белизна лепестка обусловлена присутствием системы межклетных ходов, заполненных воздухом, рассеивающих падающий на них свет. В этом отношении мезофилл лепестка подобен матовому стеклу, непрозрачность и белизна которого создается шероховатой поверхностью, сильно рассеивающей свет. Если белоснежный лепесток выдержать в воде, чтобы межклетники его пропитались водой, то лепестки теряют белизну и становятся почти прозрачными.
Пигменты лепестков бывают двух типов: одни растворены в клеточном соке, другие относятся к пигментам пластид. Пигменты клеточного сока — антациан и антохлор — в лепестках распространены преимущественно в эпидермальных клетках. Антохлор и антоциан встречаются не только в лепестках цветков одного и того же вида растений или в лепестках одного и того же цветка, но даже в клетках одного и того же лепестка или в одной и той же клетке. Между антоцианом и антохлором существует родственная химическая связь: и тот и другой имеют глюкозидный характер. Иногда в одной и той же эпидермальной клетке лепестка сосочек клетки занят антохлором, а базальная ее часть антоцианом или наоборот (как в цветках львиного зева). Оба пигмента нередко встречаются в смеси друг с другом. Такие участки со смешанной пигментацией клеток встречаются в местах перехода от антоциановой к антохлоровой окраске.
Пигменты клеточного сока не всегда занимают все эпидермальные клетки лепестков. Часто пигмент распространен неравномерно, местами сгущаясь в одних и ослабляясь или даже пропадая в других участках лепестка. Нередко в базальной части лепестков, в том месте их, где образуется трубка венчика, пигментация лепестков сильно ослабляется или же полностью отсутствует. На отгибе лепестков, наоборот, пигментация может быть, очень интенсивной. В этих участках лепестков только над жилками интенсивность пигментации заметно ослабляется или же меняется окраска пигмента. Например, у синих лепестков бывают красноватые жилки.
Сосочки, а также чередование густопигментированных клеток с клетками слабопигментированными или же полностью лишенными пигмента создают громадное разнообразие в характере окраски лепестков, помимо изменения окраски самого пигмента. Усилению разнообразия способствуют также различные выросты, изменяющие очертания поверхности лепестка и нередко окрашенные несколько иначе, чем прочая часть пластинки лепестка.
Из пластид в лепестках доминирующее значение имеют хромопласты, придающие лепесткам различную окраску разнообразнейших тонов, от бледно-желтого до красноватого или оранжевого. Иногда преимущественно в лепестках, относительно долго сохраняющихся в цветке, строма хромопласта разрушается, пигмент или кристаллизуется, или образует маслянистые капли.
Хромопласты лепестков расположены преимущественно в эпидермальных клетках.
Зеленые пластиды в лепестках образуются как в мезофилле, так и в эпидермальных клетках, при этом они никогда не имеют тех размеров и той ярко-зеленой окраски, какие свойственны хлоропластам листьев; они мельче и желтоватого оттенка. По-видимому, хлоропласты в лепестках, даже вполне сформировавшихся и совершенно белых при рассматривании невооруженным глазом, всегда присутствуют. Некоторое количество бледно-зеленых пластид сохраняется в клетках мезофилла, расположенных в непосредственном соприкосновении с жилками. Хлоропласты наиболее хорошо выражены в молодых лепестках, особенно в лепестках цветковых бутонов. В лепестках только что образовавшихся бутонов хлоропласты крупные и ярко-зеленые. Затем по мере дальнейшего развития цветка часть хлоропластов разрушается, часть же заметно деградирует в своих размерах и пигментации.
Цветоножка у различных растений построена сходно, напоминая в некоторых отношениях структуру молодого стебля. Если цветоножка прямостоячая, то все сосудистые пучки ее одинаковы и расположены кольцом. Если же цветок свисает и цветоножка отходит сбоку стебля или отогнута, то сосудистые пучки развиваются неравномерно. В одних пучках определенно выявлены ксилема и флоэма, другие содержат незначительное количество дифференцированных элементов ксилемы и флоэмы.
При превращении цветоножки в плодоножку все живые клетки разрастаются, причем происходит сильное одревеснение большинства тканей, особенно тканей центрального цилиндра.
В молодой плодоножке, начинающей развиваться после оплодотворения и несущей только что начавший формироваться плод, в первую очередь дифференцируется твердый луб, элементы ксилемы сильно разрастаются, клетки сердцевинных лучей одревесневают. В ксилеме плодоножки образуется очень мало широкополостных сосудов; она почти целиком состоит из либриформа и трахеидоподобных анатомических элементов с толстыми оболочками.
Во вполне зрелой плодоножке одревеснение слагающих ее анатомических элементов распространяется также на значительную часть сердцевины. При такой структуре плодоножка представляет собой весьма мощное, с механической точки зрения, образование, приспособленное выдерживать значительную нагрузку, и в то же время достаточно гибкое.
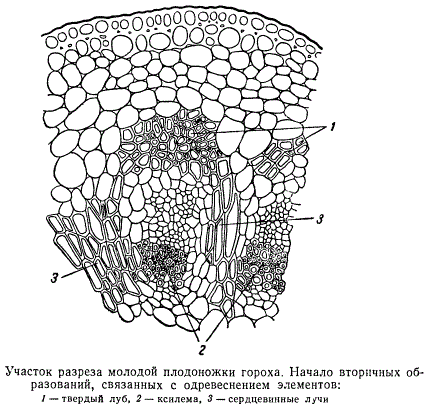
Камбия в сосудистых пучках цветоножки и плодоножки не имеется; увеличение размеров поперечника при превращении цветоножки в плодоножку, по-видимому, обусловлено одним лишь разрастанием и дальнейшей дифференцировкой существовавших в цветоножке полумеристематических клеток. Следовательно, все возникающие в цветоножке элементы каждого сосудисто-волокнистого пучка первичного (не камбиогенного) происхождения. Спиральные проводящие элементы ксилемы проводящих пучков цветоножки в плодоножках постепенно облитерируются и разрушаются.
В зависимости от свойств растения, плодоножки в деталях могут быть построены различно. У плодовых культур с объемистыми и тяжелыми плодами в плодоножках, кроме сильного общего одревеснения клеток, в добавление к твердому лубу дифференцируются еще специальные механические элементы в виде различного рода склереид.
У некоторых травянистых растений, имеющих большие и тяжелые плоды, при созревании свисающие или лежащие на земле, как, например, у тыквенных, одревеснение оболочек анатомических элементов сосудистых пучков в плодоножках также бывает весьма значительным, хотя склеренхиматизация тканей выражена не столь сильно, как в плодоножках плодовых; в плодоножках тыквенных основная паренхима одревесневает относительно слабо.
После окончательного созревания плодов в плодоножках, как и в черешках листьев, обычно развивается разделительная ткань и плод отпадает. Однако у многих растений, особенно травянистых, плодоножки просто высыхают и, становясь хрупкими, легко ломаются даже при незначительных механических воздействиях на них, например при ветре.