В клетках как низших, так и высших растений можно наблюдать передвижение и переориентацию хлоропластов, хромопласты и лейкопласты лишены такой особенности. Весьма показательно, что разрушение зеленых пигментов приводит к потере способности хлоропластов передвигаться, что происходит при пожелтении листьев осенью, а также при длительном выдерживании растения в темноте. Способность к движению может возобновляться, например, в том случае, если происходит позеленение этиолированных листьев в условиях хорошего освещения. Причем хлоропласта начинают передвигаться только тогда, когда восстанавливается их фотосинтетическая деятельность.
Вышеприведенные факты наталкивают на мысль о том, что движение пластид тесно связано с процессом фотосинтеза. Изменение расположения хлоропластов в клетке, вызываемое их движением и переориентацией, следует рассматривать как приспособление растений, с помощью которого обеспечиваются наиболее благоприятные условия для работы фотосинтетического аппарата. В первую очередь это относится к условиям освещения, поскольку на расположение пластид и их ориентацию в клетке свет обычно влияет сильнее, чем другие факторы внешней среды. Количество поглощенного листом света во многом зависит не только от общего числа и размеров пластид, но и от их расположения в пределах клеток.
Типы расположения хлоропластов
В зависимости от условий освещения у высших растений различают три основных типа расположения хлоропластов в клетке: эпистрофию, парастрофию и апострофию.
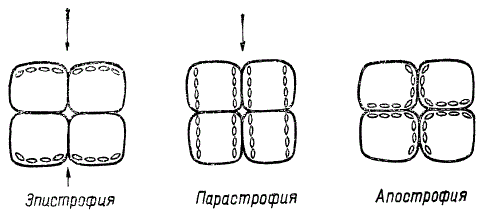
Схема расположения хлоропластов в клетках листа. Стрелки показывают направление света
Эпистрофия характерна для условий слабого освещения. Под влиянием света низких интенсивностей хлоропласты совершают движение к наружной стенке клеточной оболочки и ориентируются вдоль нее перпендикулярно направлению падающих лучей, что способствует максимальному поглощению световой энергии.
Под парастрофией понимается такое расположение хлоропластов вдоль боковых стенок клеточной оболочки, которое вызвано интенсивным освещением. Причем эти органоиды не только отходят к боковым стенкам, но и поворачиваются ребром, т. е. оказываются параллельными падающему свету. В таком положении они освещаются рассеянным светом и поглощают меньше световой энергии, а главное — защищены от прямой солнечной радиации, особенно от вредного действия коротковолновых лучей.
Ночью наблюдается апострофное расположение, когда хлоропласты находятся и вдоль боковых стенок, и вдоль внутренней стенки клеточной оболочки. Иногда в темноте они могут группироваться вокруг ядра либо беспорядочно рассеиваться по всей цитоплазме.
Передвижение пластид
Передвижение пластид можно изучать с помощью непосредственного микроскопического наблюдения, регистрируя их положение в клетке через определенные промежутки времени. Кроме того, в настоящее время для этих целей используется микрокиносъемка с последующим анализом фильмов. Посредством этих методов было установлено, что скорость и путь движения хлоропластов изменяются довольно значительно не только у растений разных видов, но даже в пределах одной и той же клетки.
У мхов и некоторых других растений хлоропласта движутся с довольно постоянной скоростью, вплоть до установления окончательного расположения, проделывая относительно прямой или слегка волнистый путь. У растений другой группы, к которой можно отнести элодею и валлиснерию, пластиды непрерывно изменяют направление движения, вычерчивая сложный путь с многочисленными изгибами и петлями. Скорость их движения также очень сильно меняется. Некоторые растения, например ряска, занимают промежуточное положение между этими двумя крайними типами.
В качестве примера, иллюстрирующего степень изменения скорости движения хлоропластов, приведем некоторые данные, полученные польскими учеными А. Зужицкой и Я. Зужицким. Средняя скорость их движения, вызванного действием сильного света, равняется 5,4 мкм/сек у ряски тройчатой (Lemna trisulca) и 25 мкм/сек у элодеи (Elodea densa). Кратковременная максимальная скорость движения отдельных хлоропластов у этих же растений может достигать соответственно 26,4 мкм/сек и 336 мкм/сек.
У растений разных видов передвижение хлоропластов к наружным или к боковым стенкам клеточной оболочки вызывается светом неодинаковой интенсивности. Особенно большие различия наблюдаются между светолюбивыми и теневыносливыми растениями. Переход от эпистрофии к парастрофии происходит у светолюбивых растений при более высоких интенсивностях света, чем у теневыносливых. Даже у представителей одного и того же вида, но произрастающих в условиях неодинакового освещения, эти органоиды различаются по своей чувствительности к интенсивности света. У листьев, адаптированных к более яркому освещению, переход от эпистрофии к парастрофии также происходит под влиянием света большей интенсивности.
До сих пор нет единой точки зрения на то, как влияет свет разной длины волны на перемещение пластид. Большинство исследователей считает, что только сине-фиолетовые лучи видимого участка спектра и близкий ультрафиолет оказывают эффективное действие на хлоропласты и вызывают их движение.
Изменение расположения пластид в клетке отражается на их работоспособности, а следовательно, и на интенсивности фотосинтеза. В опытах по изучению этого вопроса сначала достигалась либо эпистрофия, либо парастрофия, либо апострофия путем выдерживания растений при соответствующих условиях освещения. Затем у листьев, отличающихся типом расположения хлоропластов, определялась интенсивность фотосинтеза в одинаковых условиях. Опыты, проведенные по такой схеме, показали, что при освещении ярким светом максимум интенсивности фотосинтеза получается в случае парастрофного расположения, которое обеспечивает наиболее благоприятные для фотосинтеза условия освещения.
При слабом освещении наибольшая интенсивность фотосинтеза обнаруживается лишь при эпистрофии.
Передвижение и расположение пластид обусловливаются действием не только света, но и некоторых химических веществ, в частности СО2. Уже давно было подмечено, что в отсутствие углекислого газа хлоропласта не способны передвигаться под влиянием интенсивного освещения к боковым стенкам клеточной оболочки, но стоит лишь такие клетки снабдить углекислотой, как начинается движение пластид в положение парастрофии.
Движение хлоропластов может совершаться только в определенном интервале температур, который несколько отличается у разных видов растений. В качестве примера укажем, что у мужоции (Mougeotia spec.) движение пластид наблюдается в пределах от 1 до 30° С, а у селагинеллы (Selaginella martensii) — при температурах не ниже 5 и не выше 35° С. Повышение теплоустойчивости растений путем тепловой закалки приводит к тому, что передвижение приостанавливается при более высокой температуре, чем обычно.
Движение хлоропластов представляет собой одну из их естественных функций, характеризующих нормальное физиологическое состояние клетки и ее жизнеспособность. Подавление функции движения указывает на повреждение клетки и нарушение ее жизнедеятельности. Исходя именно из таких представлений, А. Г. Ломакин предложил использовать перемещение пластид, в частности переход от эпистрофии к парастрофии при интенсивном освещении, в качестве критерия устойчивости клетки к высоким температурам и другим повреждающим агентам. С помощью такого критерия можно определять также эффективность метода тепловой закалки и следить за процессом восстановления поврежденных клеток листа в результате нагрева или другого воздействия.
Интересное явление суточной ритмичности в движении хлоропластов в пределах клетки даже при константных условиях внешней среды обнаружили И. И. Гунар и И. И. Богачева. Растения выращивались в камере искусственного климата при постоянной температуре (20° С) и постоянной интенсивности света (около 5000 лк). Ежесуточно растения находились 12 часов на свету и 12 часов в темноте. При таком режиме хлоропласта клеток листа совершали в течение суток определенные ритмичные перемещения: днем они находились в положении полной парастрофии, вечером, еще до выключения света, начинали двигаться к внутренним стенкам клеточной оболочки, а ночью переходили в апострофное положение. Утром снова совершалось их перемещение к боковым стенкам, причем переход к парастрофии начинался еще до включения света, которое производилось в 8 часов утра. При перенесении этих растений в условия непрерывного освещения указанная ритмичность сохранялась в течение двух суток, несмотря на то что свет не прерывался темнотой.
О механизме движения хлоропластов
У большинства растений хлоропласта взвешены в цитоплазме таким образом, что обладают свободой движения. Они могут перемещаться в пределах клетки вполне самостоятельно и активно, а не только переноситься пассивно вместе с потоком цитоплазмы.
Несмотря на усиленное изучение движения пластид, сущность этого явления до сих пор во многом остается не расшифрованной. Все предложенные теории весьма гипотетичны и вскрывают далеко не полно механизм движений. В качестве примера очень кратко остановимся на некоторых из них.
Согласно одной из наиболее распространенных гипотез пластиды осуществляют активные движения, подобно амебам, путем образования псевдоподий (временных выступов периферической части пластид) и перетекания содержимого в этом направлении. Недостатки гипотезы амебоидных движений можно видеть в том, что иногда нельзя вскрыть прямую зависимость между направлением движения и образованием псевдоподий, а также в том, что у многих видов растений псевдоподии вообще не удается обнаружить.
По мнению других исследователей, первостепенное значение для движений имеют силы поверхностного натяжения на границе хлоропласта с цитоплазмой. Различия в этих силах на разных сторонах пластиды и обусловливают ее перемещение.
Согласно электрокинетической гипотезе притяжение пластид к стенкам клеточной оболочки или, напротив, отталкивание их, а также различные перемещения могут вызываться изменениями знака их электрических зарядов. Обоснованием этого может служить давно установленный факт изменения электрического заряда пластид под влиянием условий освещения. Пока неясно, достаточно ли для перемещения пластид одних только сил, возникающих при изменении поверхностного натяжения либо электрических зарядов.
Результаты недавно выполненных исследований указывают на то, что движения хлоропластов, как и цитоплазмы, тесно связаны с обменом веществ клетки. Важную роль в превращении химической энергии, которая высвобождается при обмене веществ, в механическую энергию движения, по-видимому, играют соединения с макроэргическими связями, особенно АТФ.
Источник: Н.Н. Овчинников, Н.М. Шиханова. Фотосинтез. Пособие для учителей. Изд-во «Просвещение». Москва. 1972