Проявление действия гена имеет определенные характеристики.
Один и тот же мутантный ген у разных организмов может проявить свой эффект различным образом. Это обусловлено генотипом данного организма и условиями внешней среды, при которых протекает его онтогенез. Фенотипическое проявление гена может варьировать по степени выраженности признака. Это явление Н. В. Тимофеев-Ресовский еще в 1927 г. предложил называть экспрессивностью гена. Действие гена может быть более или менее константным, стойким в своем проявлении или нестойким, вариабильным. С изменчивостью проявления мутантного гена у разных организмов мы действительно встречаемся довольно часто. У дрозофилы имеется «безглазая» мутантная форма (eyeless) с сильно редуцированным числом фасеток. Просматривая потомство одной родительской пары, можно видеть, что у одних мух глаза почти полностью лишены фасеток, тогда как у других число фасеток в глазах достигает половины нормального числа. Такое же явление наблюдается в реализации многих признаков и у других животных и растений.
Один и тот же мутантный признак может проявляться у одних и не проявляться у других особей родственной группы. Это явление Н. В. Тимофеев-Ресовский назвал пенетрантностью проявления гена. Пенетрантность измеряется по проценту особей в популяции, имеющих мутантный фенотип. При полной пенетрантности (100%) мутантный ген проявляет свое действие у каждой особи, обладающей им; при неполной пенетрантности (меньше 100%) ген проявляет свой фенотипический эффект не у всех особей.
Экспрессивность, как и пенетрантность, обусловлена взаимодействием генов в генотипе и различной реакцией последнего на факторы внешней среды. Экспрессивность и пенетрантность характеризуют фенотипическое проявление гена. Пенетрантность отражает гетерогенность линий, популяций не по основному гену, определяющему конкретный признак, а по генам — модификаторам, создающим генотипическую среду для проявления гена. Экспрессивность есть реакция сходных генотипов на среду. Оба указанных явления могут иметь приспособительное значение для жизни организма и популяции, и поэтому экспрессивность и пенетрантность проявления гена поддерживаются естественным отбором. Эти два явления очень важно учитывать и при искусственном отборе.
Экспрессивность гена в развитии зависит от действия факторов внешней среды. Легче всего пока удается проследить влияние различных внешних агентов на мутантные гены. Так, у кукурузы известны мутантные гены, определяющие карликовость растений, положительный геотропизм (наклоняющиеся растения) и др. В основе действия этих генов лежат соответствующие биохимические изменения. Известно, например, что для нормального роста растения необходимы ростовые вещества типа ауксинов. У мутантной карликовой формы кукурузы ауксин вырабатывается нормально, но ген карликовости тормозит образование фермента, который окисляет ауксин, вследствие чего понижена активность ауксина, что и приводит к торможению роста растений. Если на такое растение воздействовать во время роста гиббереллиновой кислотой, то растение ускоряет рост и становится по фенотипу неотличимым от нормального. Добавка гиббереллиновой кислоты как бы восполняет то, что должна была бы произвести нормальная аллель гена карликовости.
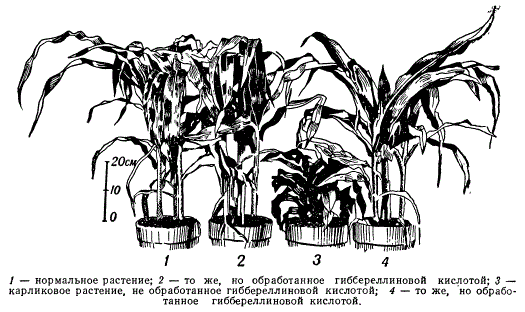
Влияние гиббереллиновой кислоты на рост кукурузы
Из этого примера видно, что ген контролирует образование определенного фермента, который изменяет характер роста растения. Таким образом, зная механизм действия мутантного гена, можно исправлять и нормализовывать вызываемые им дефекты.
Вспомним, что гималайская окраска кролика определяется одним членом серии множественных аллелей — с11. Обычное фенотипическое проявление этого гена при нормальной температуре (около 20°) характеризуется тем, что при общей белой окраске шерсти кончики лап, уши, нос и хвост кролика оказываются черными.

Фенотипическое изменение окраски шерсти гималайского кролика под влиянием различных температур
Такая окраска зависит как от определенных биохимических реакций, протекающих в коже, связанных с выработкой меланистических пигментов, так и от температуры окружающей среды. На том же рисунке показано, что кролик, выращенный при температуре выше 30°, оказывается сплошь белым. Если же выщипать небольшой участок белой шерсти и потом систематически его охлаждать, то на нем вырастает черная шерсть. В данном случае действие температуры сказывается на проявлении гена, влияя на выработку определенных ферментов.
У растения примулы известен ген окраски цветка, который также проявляет свое действие в зависимости от температуры. Если растения выращиваются при температуре 30—35° и высокой влажности, то цветки будут белыми, а при более низкой температуре — красными.
Еще в 1935 г. Ф. А. Смирнов провел работу по изучению числа индуцированных мутаций у дрозофилы: летальных, семилетальных и мутаций с повышенной и нормальной жизнеспособностью, и обнаружил разное соотношение перечисленных классов в разных температурных условиях. Позднее это же подтвердилось и на популяциях Drosophila pseudoobscura. Из дикой популяции этого вида были выделены мутанты, которые нормально развивались при температуре 16,5°, при 21° они были полулегальными, а при 25° оказывались полностью летальными. Такого рода исследования теперь ведутся на мутациях микроорганизмов. Эти мутации называются amber-мутациями.
У наездника Habrobracon hebitor известен ген kidney (k). Он имеет почти 100%-ную пенетрантность как леталь при 30°, а при низкой температуре развития почти не проявляется. Такого типа зависимость пенетрантности от условий среды известна для большинства мутаций у всех животных, растений и микроорганизмов.
Действие одного и того же фактора внешней среды сказывается различным образом на разных генах, и разные факторы различным образом влияют на проявление одного и того же гена. Изучение влияния факторов внешней среды показало, что некоторые рецессивные гены, которые в обычных условиях в гетерозиготном состоянии фенотипически не проявляются, могут проявиться при измененных условиях.