Как известно, доминирование есть подавление действия одной аллели другой аллелью, представляющих собой один ген: А > а, В > b, С > с и т. д.
Но существует взаимодействие, при котором аллель одного из генов подавляет действие аллелей других генов, например А > В или В А, а > В или b > А и т. д. Такое явление «доминирования» между генами называется эпистазом.
Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию.
Гены, подавляющие действие других генов, называются супрессорами, или ингибиторами. Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных (млекопитающие, птицы, насекомые) и у растений. Обычно они обозначаются I или Su в случае доминантного состояния генов и i или su для их рецессивных аллелей (от английских слов inhibitor или supressor).
В настоящее время эпистаз делят на два типа: доминантный и рецессивный.
Под доминантным эпистазом понимают подавление доминантной аллелью одного гена действия аллельной пары другого гена. Из многих примеров доминантного эпистаза, установленных как на животных, так и на растениях, приведем лишь отдельные.
Некоторые породы кур имеют белое оперение (белый леггорн, белый примутрок и др.), другие же породы имеют окрашенное оперение (австралорп, ньюгемпшир, полосатый плимутрок и др.). Белое оперение разных пород кур определяется несколькими различными генами. Так, например, доминантная белая окраска определяется генами CCII (белые леггорны), а рецессивная белая — ccii (белые суссексы, белые минорки, белые плимутроки). Ген С определяет наличие предшественник пигменту (хромогена), т. е. окрашенность пера, его аллель с — отсутствие хромогена и, следовательно, неокрашенность пера птицы. Ген I является подавителем действия гена С, аллель i не подавляет его действия. В присутствии даже одной дозы гена I в генотипе птицы действие генов окраски не проявится. Поэтому при скрещивании белых леггорнов CCII с цветными породами CCii, как правило, доминирует белая окраска CCIi. При скрещивании белых плимутроков ccii с окрашенными породами CCii гибриды оказываются окрашенными Ccii. Следовательно, у леггорнов белая окраска является доминантной, а у плимутроков — рецессивной.
Если же производится скрещивание белых леггорнов CCII с рецессивными белыми плимутроками ccii, то в первом поколении цыплята оказываются тоже белыми CcIi. При скрещивании между Собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13/16 белых : 3/16 окрашенных.
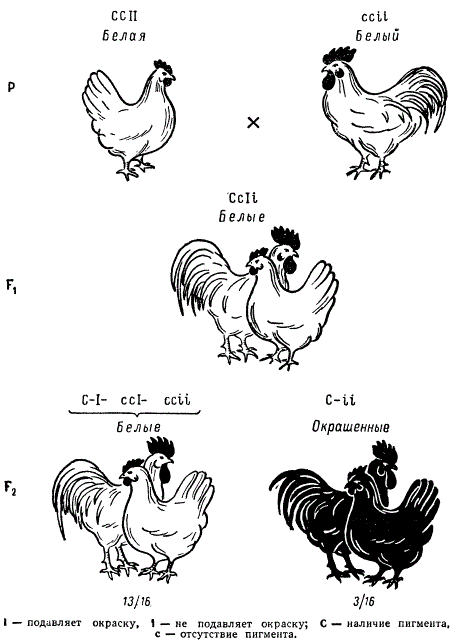
Наследование окраски у кур при взаимодействии двух пар генов (эпистаз)
Как можно объяснить полученное отношение? Прежде всего, данное отношение говорит о расщеплении по двум генам, это расщепление можно представить как 9(С—I—) + 3(ccI—) + 1(ccii) = 13 и 3(С—ii), что соответствует формуле 9:3:3:1.
Очевидно, в этом случае окраска леггорнов обязана не присутствию особых генов белой окраски, а действию гена — подавителя окраски (I—). Тогда генотип гомозиготных белых леггорнов должен быть ССII, где I является геном — подавителем окраски, а С — геном окраски. Белые плимутроки по генотипу должны быть гомозиготными по двум рецессивным факторам ccii, где с — отсутствие окраски и i — отсутствие подавления окраски. В силу эпистатирования I > С гибридные куры первого поколения CcIi должны быть белыми. В F2 все куры с генотипами 9/16 С—I—, 3/16 ccI— и 1/16 ccii также должны быть белыми, и лишь куры одного фенотипического класса 3/16 (С—ii) оказываются окрашенными, поскольку здесь содержится ген окраски и нет его подавителя.
Таким образом, подавление действия доминантной аллели гена, Определяющего развитие окраски, доминантной аллелью другого гена (подавителем) обусловливает в F2 расщепление по фенотипу в отношении 13 : 3.
Доминантный эпистаз может давать и другое отношение при расщеплении в F2 по фенотипу, а именно 12 : 3 : 1. В этом случае форма гомозиготная по обоим рецессивным факторам aabb, будет фенотипически отличима от форм с доминантными аллелями двух генов А—В— и форм с одной из них: ааВ— и А—bb. Такое расцепление установлено для наследования окраски плодов у тыквы, кожуры у лука и других признаков. В этом случае в расщеплении также принимает участие доминантный ингибитор.
Мы разобрали взаимодействие только двух генов. В действительности по типу эпистаза взаимодействуют многие гены. Гены-подавители обычно не определяют сами какой-либо качественной реакции или синтетического процесса, а лишь подавляют действие других генов. Однако когда мы говорим, что ген-подавитель не имеет своего качественного влияния на признак, то это относится только к данному признаку. На самом же деле ингибитор, подавляя, например, пигментообразование, может оказывать плейотропное действие на другие свойства и признаки.
Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивная аллель одного гена, будучи в гомозиготном состоянии, не дает возможности проявиться доминантной или рецессивной аллелям других генов: аа > В или aa > bb.
Мы уже имели случай познакомиться с расщеплением 9:3:4 как результатом комплементарного взаимодействия генов. Но эти же случаи можно рассматривать и как пример рецессивного эпистаза.
При скрещивании черных мышей (ААbb) с белыми (ааВВ) все особи F1 (АаВb) имеют окраску типа агути, а в F2 9/16 всех особей сказываются агути (А—В—), 3/16 черные (A—bb) и 4/16 белые (ааВ— и aabb). Эти результаты можно объяснить, предположив, что имеет место рецессивный эпистаз типа ааВ—. При этом мыши генотипа ааВ— оказываются белыми потому, что ген а в гомозиготном состоянии, обусловливая отсутствие пигмента, препятствует тем самым проявлению гена-распределителя пигмента В.
Кроме описанных случаев одинарного рецессивного эпистаза, действуют и такие, когда рецессивная аллель каждого из генов в гомозиготном состоянии одновременно реципрокно подавляет действием доминантных аллелей каждого из генов, т. е. аа эпистатирует над В—, a bb над А—. Такое взаимодействие двух подавляющих рецессивных генов называют двойным рецессивным эпистазом. При этом в дигибридном скрещивании расщепление по фенотипу будет соответствовать 9 : 7, как и в случае комплементарного взаимодействия генов.
Следовательно, одно и то же отношение можно трактовать и как комплементарное взаимодействие, и как эпистатирование. Сам по себе генетический анализ наследования при взаимодействии генов без учета биохимии и физиологии развития признака в онтогенезе не может раскрыть природы этого взаимодействия. Но без генетического анализа нельзя понять наследственной детерминации развития этих признаков.