Эволюция организмов и есть непрерывная замена одних генотипов другими.
Эта смена происходит в популяции путем изменения численного соотношения качественно различающихся генотипов. Изменение соотношения генотипов составляет сущность динамики генетической структуры популяции. Генетическая изменчивость популяции складывается, как мы уже говорили, из мутационной и комбинативной изменчивости.
Равновесие генотипов в популяции, основанное на сохранении относительных частот генов, изменяется под влиянием ряда постоянно действующих факторов, к которым относятся: 1) мутационная изменчивость, 2) действие отбора, 3) миграция, 4) изменение численности популяции, 5) избирательность спаривания и оплодотворения и ряд других факторов. Рассмотрим кратко некоторые из них.
Роль мутационного процесса в динамике популяции
Сохранение даже относительного постоянства частоты генов в популяции возможно при допущении, что они не мутируют.
Однако известно, что это не так. Мутации составляют первичный источник наследственной изменчивости в эволюции; и хотя каждый ген спонтанно мутирует довольно редко, общее количество различных мутаций может быть достаточно большим, поскольку число генов бесконечно велико.
Если аллель А мутирует с определенной скоростью в а, то очевидно, что в генофонде популяции постепенно в ряду поколений частота аллели А будет уменьшаться на число измененных генов, а-частота аллели а возрастает на эту же величину. Это, естественно, приведет к нарушению равновесия в популяции, к изменению ее генетического состава. В каждом поколении генофонд может пополняться значительным числом мутаций как одного определенного локуса, так и разных. Этот процесс называют мутационным давлением. Мутационное давление играет важную роль в генетической динамике популяции.
Изменение гена может происходить в прямом и обратном направлениях и что каждый ген, по-видимому, имеет определенную скорость прямого и обратного мутирования. Следовательно, частота аллелей разных генов в популяции будет изменяться в зависимости от мутационного давления, т. е. от соотношения прямых и обратных мутаций: Особенно существенным это мутационное давление оказывается в отношении так называемых высокомутабильных, т. е. часто мутирующих, генов.
Однако распространение новой мутации в генофонде популяции зависит еще и от того, в какой мере она влияет на жизнеспособность и плодовитость особи. Мы знаем, что существуют летальные, полулетальные, видимые (морфологические) и биохимические мутации, вызывающие различные крупные или мелкие изменения. Некоторые генетики признают наличие так называемых нейтральных мутаций, не затрагивающих жизнеспособность организма. Мы считаем, что таких мутаций не существует. Лишь неполнота наших знаний
о действии мутантных аллелей ведет к выделению мутаций такого типа. На самом деле мутации всегда или дают преимущество их носителям, или наносят им вред вплоть до вызывания летального эффекта.
Так же невозможно на данном уровне знаний во всех случаях с определенностью говорить о «мелких» и «крупных» мутациях и степени их ценности в процессе отбора, ибо мы еще мало знаем о плейотропном действии многих из них. Мутации, вредные в гомозиготном состоянии, могут оказываться полезными в гетерозиготном состоянии. Так, например, рецессивный ген, обусловливающий серповидность эритроцитов у человека, вреден в гомозиготном состоянии, а в гетерозиготном — повышает устойчивость к малярии.
Всякая вновь возникшая мутация вызывает изменение целостной генетической системы, стабилизировавшейся в процессе исторического развития. Поэтому мутация, не прошедшая отбора в системе генотипа, нарушает исторически сложившиеся корреляции функций организма в индивидуальном развитии. В этом смысле каждая отдельная новая мутация в подавляющем большинстве случаев вначале оказывается вредной и лишь очень редко сразу может оказаться полезной для вида, имея некоторое положительное селективное значение.
Доминантная мутация подвергается контролю со стороны отбора уже в гетерозиготе. Рецессивная мутация может подвергаться отбору лишь в том случае, если она размножится в популяции до определенного уровня и перейдет в гомозиготное состояние. Размножение рецессивной мутации в популяции зависит 1) от характера проявления мутантного гена в гомозиготном состоянии, 2) от частоты встречаемости гетерозиготных по данному гену особей. Чем меньше размер популяции, тем вероятнее скрещивание гетерозиготных особей и, напротив, чем больше численность популяции, тем меньше вероятность проявления рецессивного гена в гомозиготном состоянии (при условии одинаковой пропорции гетерозигот в популяции). В большой популяции потребуется более длительный срок на размножение мутации. Таким образом, распространение мутации в популяции зависит и от ее численности. Как только начнут появляться мутантные фенотипы, они подвергнутся контролю со стороны отбора, хотя следует подчеркнуть, что некоторые рецессивные мутации могут контролироваться отбором еще в гетерозиготном состоянии.
Роль отбора в генетической динамике популяций
Отбором называют процесс переживания организмов, генотипы которых обеспечивают им наибольшую приспособляемость к условиям среды в течение индивидуальной жизни и которые вследствие этого оставляют наибольшее число потомков.
Отбор обычно разделяют на искусственный и естественный. В принципе оба типа отбора не различаются, но каждый из них имеет свои важные особенности.
Прежде всего, искусственный отбор производится человеком с целью создания пород животных, сортов растений и штаммов микроорганизмов для повышения производительных сил общества, а естественный отбор осуществляется в естественных, природных условиях жизни организмов под влиянием факторов среды.
Другим различием между этими двумя типами отбора является то, что человек, опираясь на знание законов наследственности и изменчивости, может отобрать нужные ему формы в более короткий срок in обеспечить больший коэффициент их размножения, чем это происходит в природе. При искусственном отборе человек, во-первых, ограничивает случайное сочетание всех возможных генотипов, которое здесь он чаще всего заменяет подбором родительских пар с нужными генотипами, и, во-вторых, он может искусственно увеличивать мутационную изменчивость. В природных популяциях сочетание генотипов при скрещивании осуществляется случайно. Эта случайность оказывается закономерной необходимостью в развитии органической природы.
Вероятность того, что организм выживет и даст потомство, зависит от степени его приспособленности к среде. Чем более надежными приспособительными механизмами обладают организмы, тем вероятнее их сохранение и процветание популяции. Знание генетики популяции позволяет определять селективную ценность генотипов.
Допустим, что особи, гомозиготные по рецессивной аллели гена (аа), дают 99 потомков на каждые 100 потомков, производимых особями с доминантными генами (АА и Да). Селективную ценность последних можно принять за 1,00, тогда таковая рецессивных гомозигот составит 0,99. Разность этих величин будет выражать коэффициент отбора генотипов — S, в данном случае S=1,00-0,99=0,01.
Если определенные генотипы обусловливают равную выживаемость и плодовитость особей, то коэффициент отбора в этом случае будет равен 0. Если же один из генотипов вызывает обязательную гибель или полную стерильность, то коэффициент отбора окажется равен 1. Очевидно, что особи, несущие соответствующие аллели в гомозиготном состоянии (если они рецессивны), будут выбраковываться отбором, и частота этого гена в поколениях панмиктической популяции сократится. Следовательно, отбор ограничивает распространение неблагоприятных генов. При таком понимании борьба за существование выглядит как «соревнование» между организмами за передачу наследственных свойств потомкам. Концентрация более вредных мутаций в популяции снижается в поколениях скорее, чем менее вредных. И наоборот, концентрация мутаций, имеющих существенное приспособительное значение, будет возрастать быстрее, чем мутаций, менее полезных.
Очевидно также, что концентрация генов определяется еще и численностью популяции. В малых популяциях, как мы говорили, увеличивается вероятность встречи гамет с одинаковыми аллелями того или иного гена, что приводит к повышению частоты возникновения гомозиготных форм, и отбор скорее начинает устранять вредные гены и накапливать полезные. Но одновременное этим в ограниченной популяции увеличивается момент случайности в накоплении отдельных генотипов. При сокращении численности популяции по каким-то случайно сложившимся обстоятельствам в ней могут сохраняться одни мутантные гены, а другие также случайно элиминируются. При последующем увеличении численности популяции частота этих случайно сохранившихся генов может довольно быстро возрасти. Это явление называют генетическим дрифтом.
Скорость устранения доминантных и рецессивных аллелей из популяции различна. Доминантные мутации так же, как и рецессивные, могут быть летальными, полулетальными, определять полную или частичную стерильность, вызывать различные морфологические изменения и физиологическую (биохимическую) недостаточность. Они могут быть с полной или частичной проявляемостью (пенетрантностью) и различной экспрессивностью. Ясно, что организмы, несущие летальные доминантные гены или гены стерильности с полным проявлением, устраняются отбором в первом же поколении. Все другие доминантные гены, лишь снижающие жизнеспособность или сокращающие плодовитость, и даже летальные гены, но с неполной пенетрантностью, могут сохраняться в популяции некоторое время и лишь постепенно элиминируются в ряду поколений. Однако полной элиминации этих генов будет противодействовать процесс мутирования — повторного возникновения тех же мутаций. В случае, когда доминантные гены имеют приспособительную ценность, они подхватываются отбором, и их частота в популяции быстро возрастает. Иначе говоря, доминантный ген в каждом поколении находится под контролем отбора.
Рецессивные мутации, в отличие от доминантных, могут находиться в популяции в скрытом, гетерозиготном, состоянии, накапливаться в ней, создавая огромный мутационный резерв. Чем меньше частота рецессивных аллелей в популяции, тем в большей степени гетерозиготы превосходят в численном отношении рецессивные гомозиготы. Это вытекает из формулы Гарди — Вайнберга. Напомним, что число рецессивных гомозигот в популяции составляет (1 — q)2 в то время как число гетерозигот составляет 2q (1—q). И «чем больше будут устраняться отбором из популяции гомозиготы, тем больше возрастет роль гетерозигот, которые явятся поставщиками рецессивных аллелей в последующих поколениях. Следовательно, отбор рецессивных генов оказывается менее эффективным, чем доминантных.
Для иллюстрации воспользуемся следующими двумя примерами. Предположим, что рецессивные гомозиготы полностью устраняются, т. е. коэффициент отбора для них 5 = 1,00. Такой случай возможен, если гомозиготы летальны или бесплодны. Допустим, что частота такого рецессивного гена в генофонде исходной популяции равна 1 —q = 0,5. Согласно формуле Гарди — Вайнберга, такая популяция будет состоять из 25% АА, 50% Аа и 25% аа. Относительная частота гетерозигот в последующих поколениях будет значительно медленнее сокращаться в популяции, чем частота рецессивных гомозигот.
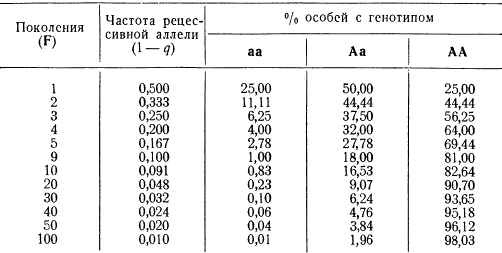
Эффективность отбора при полной элиминации рецессивных гомозигот (S=1)
Данные свидетельствуют также о том, что полное устранение из популяции рецессивных гомозигот в каждом поколении не приводит к окончательному исчезновению их даже в сотом поколении, так как гетерозиготные особи являются постоянными поставщиками гомозиготных рецессивов.
В других случаях, когда рецессивные гены определяют, например, различную степень понижения жизнеспособности, коэффициент отбора рецессивных гомозигот может быть меньше единицы. Кроме того, гены действуют в системе генотипа, поэтому в разных генотипах они могут изменять свою отборную ценность.

Эффективность отбора по рецессивному признаку в поколениях при разных коэффициентах отбора
Приводится расчет эффективности отбора в поколениях по рецессивному признаку при разных коэффициентах отбора. При этом допускается, что популяция первоначально содержала 1% гомозиготных рецессивных особей, т. е. первоначальная частота рецессивного гена была 1—q = 0,01.
Данные показывают, что при коэффициенте отбора 0,50, т. е. при элиминации из-за пониженной жизнеспособности половины особей, гомозиготных по рецессивному гену, за 20 поколений процент их появления сократится на 3/4, а при S = 0,01 за 20 поколений частота появления гомозигот уменьшается с 1 до 0,97%, т. е. всего на 0,03%.
Результаты расчетов при разных коэффициентах отбора показывают, почему даже при большом количестве поколений не могут быть быстро достигнуты сдвиги в желаемую сторону. Эти же расчеты дают представление о роли рецессивных мутаций в жизни популяции при разных условиях действия естественного отбора.
Таким образом, отбор является решающим фактором для дивергенции вида, так как он контролирует весь процесс эволюции. Сам же естественный отбор обусловлен абиотическими и биотическими факторами, составляющими внешнюю среду как для отдельного организма, так и для популяции в целом.
Факторы изоляции
Как уже отмечалось, вид слагается из отдельных генетических популяций.
Если особи одной популяции полностью или частично прекращают в силу различных причин обмениваться генами с особями других популяций, то такая популяция испытывает процесс изоляции. Если разобщение продолжается в ряду поколений, а факторы отбора в разных популяциях действуют в разных направлениях, то происходит дифференциация популяций. В последующем такие популяции могут дать начало новым разновидностям, а при более сильном расхождении — и новым видам.
Генетическая изоляция внутри вида может обеспечиваться географическими, экологическими и биологическими факторами. Любые разобщения популяции, вызванные геологическими изменениями (горообразовательные процессы, возникновение рек и водоемов), могут приводить к географической изоляции. Экологические факторы — территориально-климатические, микроклиматические, сезонно-климатические и др. — также могут создавать препятствия для свободного скрещивания организмов одного вида.
Так, например, у рыб, ведущих морской образ жизни, но возвращающихся для размножения в реки, в каждой реке и даже в отдельных притоках складывается своя особая популяция. Рыбы этих популяций могут различаться по величине, окраске во время нереста, возрастному составу, покатному возрасту молоди, времени наступления половой зрелости, количеству и размеру икринок, срокам возвращения из моря на нерест и срокам размножения.
Подобные различия имеют не только модификационную природу вследствие влияния среды, характерной для режима каждой реки, но некоторые из них наследственно обусловлены. Так, если взять оплодотворенную икру рыб из популяции, населяющей одну реку, и перенести в другую реку, населенную другой популяцией, то выросшие рыбы первой популяции и в новых условиях сохраняют характерные для них признаки. Следовательно, особенности организмов каждой из популяций наследственно закреплены и сохраняются благодаря действию отбора; направление действия отбора определяется условиями внешней среды, в которой осуществляется онтогенез.
К биологическим факторам изоляции, нарушающим свободу скрещивания, относятся генетические и физиологические. Собственно генетическими факторами изоляции являются нарушения нормального течения мейоза, препятствующие воспроизведению плодовитого потомства. Причинами таких нарушений могут быть:
- полиплоидия,
- хромосомные перестройки,
- ядерно-цитоплазматическая несовместимость,
- повышение концентрации летальных мутаций и мутаций стерильности.
Каждое из перечисленных явлений может приводить к ограничению свободного скрещивания особей и бесплодию гибридов, а следовательно, к ограничению свободного комбинирования генов, т. е. к генетической изоляции этих форм, которые внутри себя могут давать плодовитое потомство.
Изоляция популяций может обеспечиваться также и физиологическими факторами. Известно, что морские рыбы, идущие на нерест в реки, как правило, возвращаются в те реки, в которых они появились на свет и провели свою «молодость» до ската в море. Причину этого и подобных явлений часто приписывают существованию «инстинктов», т. е. врожденной способности к целенаправленным поведенческим реакциям.
С нашей точки зрения, сам процесс возврата рыб в родные реки в рассматриваемом примере не является врожденным. В его основе лежит механизм онтогенетической адаптации, а именно образование цепи условных рефлексов к факторам внешней среды при жизни мальков в реке и при скате их в море. Образуется своеобразная «программа» движения, которая при возврате «читается» в обратном порядке. Однако сама возможность такого программирования обусловлена наследственно.
Физиологический механизм, обеспечивающий возврат рыбы в свою родную реку, служит фактором изоляции для популяций рыб данной реки. Такое обособление ограничивает свободу скрещивания между особями одного и того же вида.
Условный рефлекс может участвовать в обеспечении изоляции у животных при изменении суточной и сезонной половой активности, наличия у ряда особей популяции избирательности спаривания, приводящей к нарушению панмиксии, и т. д. Так, показано, что некоторые петухи предпочтительно избирают кур определенного цвета, быки-производители охотнее покрывают одних коров, и их половой рефлекс тормозится при покрытии других. То же самое установлено для насекомых. Например, у дрозофилы серые самцы (дикого типа) в определенных условиях предпочтительно (до 75%) избирают самок желтого цвета, имеющих мутантный фенотип yellow, и значительно реже спариваются с самками черного цвета (мутация ebony). Л. З. Кайданов показал, что в основе такой избирательности лежат физиологические процессы, а именно уровень полового возбуждения самцов и образование у них полового условного рефлекса.
Обособление группы организмов из популяции увеличивает вероятность скрещивания генетически сходных особей. Изоляция, таким образом, ведет к усилению инбридинга среди аллогамных организмов. Отсюда с очевидностью следует, что сама изоляция является причиной дифференциации популяции.