Хромосомы являются неотъемлемой частью всей системы клетки. Редупликация хромосом и весь цикл деления ядра являются продуктом деятельности всей клетки.
Перекрест хромосом как сложный физиологический, физический и биохимический процесс подвержен влиянию факторов внешней среды, функционального состояния целого организма и отдельной клетки. Перекрест хромосом обусловлен и их внутренней структурой, а также их генным составом, т. е. генотипом.
Перекрест у гомогаметного и гетерогаметного полов
У большинства изученных животных и растений хиазмы и перекрест осуществляются в мейозе у обоих полов.
Кажется, не известно ни одного случая, когда бы перекрест отсутствовал у обоих полов. Но существуют отдельные виды животных, у которых мейотический кроссинговер осуществляется только у гомогаметного пола, а у гетерогаметного в норме отсутствует причем это касается не только половых хромосом, отсутствие синапсиса у которых можно было бы объяснить их гетероморфностью у гетерогаметного пола, но и аутосом.
По некоторым цитологическим данным, у самцов дрозофилы и самок шелкопряда в профазе I мейоза либо полностью отсутствует синапсис и образование хиазм, либо эти процессы происходят в профазе мейоза, а на более ранних стадиях развития половых клеток на стадии образования сперматогониев и оогониев. В данном случае осуществляется гониальный кроссинговер. У гомогаметного пола этих видов (самок дрозофилы и самцов шелкопряда) перекрест хромосом протекает нормально.
У многих видов млекопитающих, птиц, рыб и насекомых гетерогаметность пола не сказывается на процессе кроссинговера. Указания о влиянии пола на кроссинговер есть лишь для некоторых недостаточно проверенных случаев (голуби, мыши).
Влияние структуры хромосом на частоту перекреста
Как мы знаем из морфологического описания структуры хромосомы, она состоит из эухроматиновых и гетерохроматиновых участков.
Целостная структура, или индиввдуальность, хромосомы в митозе и мейозе обеспечивается наличием центромеры. Из описания роли и поведения центромер в профазе I мейоза мы знаем, что в процессе синапсиса на зиготенной стадии гомологичные хромосомы сходятся центромерными или концевыми районами. В диплотенной стадии центромеры начинают отталкиваться друг от друга. Причины притяжения и отталкивания центромер не выяснены.
Не совсем ясна и структура центромеры у разных видов организмов; является ли она одинаковой для всех или нет – мы также не знаем. Имеются лишь некоторые указания на различное строение центромер.
Схема строения центромер
Впервые центромера была описана К. Метнером в 1894 г. У абсолютного большинства организмов центромера одна и строго локализована в хромосомах. Существуют полицентрические хромосомы с несколькими центромерами, как, например, у аскариды; эти хромосомы представляют собой комплексы из многих мелких хромосом. В 1941 г. С. Пиза открыл у бразильского скорпиона диффузную центромеру. Оказалось, что такое строение центромеры характерно для целой группы Hemiptera. Такая же центромера обнаружена среди растений у ожиги (Lusula purpurea). Предполагают наличие диффузной центромеры также и у некоторых водорослей.
Локализованная центромера представляет собой довольно сложную структуру. Она состоит из нескольких мелких хромомер, у которых обнаруживается ДНК.
Центромера делится, как правило, продольно — вдоль оси хромосомы, но иногда может делиться поперечно, давая две функционирующие изохромосомы. Функция центромеры в хромосоме весьма ответственна. Центромера сохраняет целостность группы сцепления генов, обеспечивает ориентацию хромосомы в метафазе и правильное расхождение хромосом к полюсам. Если центромеру разрушить ультрафиолетовыми лучами, то хромосома оказывается ориентированной. Установлено, что около центромеры имеются, как правило, гетерохроматические районы.
Накоплены факты, которые говорят о том, что центромера с прилегающим к ней гетерохроматином играет очень важную роль я в механизме перекреста хромосом. Некоторые авторы полагают, что в метацентрических хромосомах в каждом из плеч перекрест является функцией расстояния от центромеры. В норме вблизи центромеры перекрест происходит редко. По мере удаления от центромеры частота перекреста у дрозофилы увеличивается, а затем к концам снова уменьшается. Поэтому на генетических картах дрозофилы вблизи центромеры локализуется генов больше, чем в удаленных от нее районах. У кукурузы такого влияния центромеры на кроссинговер не обнаружено.
На частоту перекреста в разных участках хромосомы влияет распределение гетерохроматиновых и эухроматиновых районов. Данные некоторых авторов указывают на то, что гетерохроматиновые участки способствуют более высокой изменчивости частоты перекреста под влиянием факторов внешней среды.
Значение гетерохроматиновых и эухроматиновых районов для кроссинговера можно рассматривать с точки зрения степени спирализации хромосомной нити в этих районах. Понижение частоты Кроссинговера в центромерном районе может быть связано с высокой спирализацией данного района хромосомы. Спирализация уменьшает цитологическое расстояние между генами, а ее усиление может препятствовать синапсису гомологичных районов и обмену.
Влияние функционального состояния организма на перекрест хромосом
Выше мы уже говорили о том, что гетерогаметного пола (самцы дрозофилы и самки тутового шелкопряда) перекрест хромосом не обнаруживается.
Однако, если эти организмы подвергнуть действию рентгеновых лучей, то в потомстве возникают кроссоверные особи. Для кукурузы установлено, что процент перекреста иногда выше В мужских спороцитах, чем в женских.
Частота кроссинговера зависит также от возраста организма (по крайней мере, это установлено для дрозофил). Если изучить перекрест между двумя генами, локализованными по генетической карте на небольшом расстоянии, скажем на расстоянии 6 морганид друг от друга, и учитывать процент кроссинговера у самок по десятидневкам откладки оплодотворенных яиц (за 1 — 10, 11—20, 21—30 дней); то частота перекреста будет колебаться соответственно 5,9, 1,8 и 3,8%. Следовательно, первый возраст соответствует максимуму, второй — спаду, а третий — подъему процента перекреста.
Есть основание полагать, что возраст у ряда организмов оказывает влияние на более общее поведение хромосом в мейозе. Установлено, например, что явление нерасхождения хромосом у дрозофилы зависит от возраста организма.
В недавнее время показано, что у женщин нерасхождение Х-хромосом в первом делении мейоза учащается в возрасте от 35 лет и старше: число детей с нерасхождением Х-хромосом, родившихся у женщин этого возраста, составляет 3,0%, а у молодых — около 0,1%. Нерасхождение хромосом часто обусловливает наследственные заболевания.
Вполне возможно влияние функционального состояния организма на течение различных стадий мейоза (а значит, и на перекрест хромосом), ибо степень спирализации хромосом, скорость прохождения различных стадий профазы может в сильной степени зависеть от физиологического состояния клеток. Это тем более вероятно, что в зависимости от физиологического состояния клеток могут изменяться соотношения химических ингредиентов хромосом и цитоплазмы, а также дисперсность коллоидов — повышаться или понижаться вязкость хромосом и окружающей кариоплазмы. Все эти факторы влияют на изменение частоты перекреста.
Влияние генотипа на перекрест хромосом
У генетика не вызывает сомнения положение о том, что все механизмы, работающие в клетке или в организме, наследственно детерминированы. Кроссинговер так же, как любой другой процесс в организме, контролируется генотипом. Подтверждением этому служит эффективность отбора линий на высокую и низкую частоту перекреста между двумя генами. Отбор линий по данному признаку указывает на роль генотипа в определении кроссинговера.
Генотип может влиять на частоту кроссинговера разными путями. На молекулярном уровне генотип регулирует точность копирования ДНК. На хромосомном уровне генотип влияет через изменение политении хромосом, степени спирализации, прочности продольных связей в нити ДНК.
На частоту перекреста в сильной степени влияют различные хромосомные перестройки, поскольку они нарушают нормальный синапсис хромосом. Имеется и межхромосомное влияние на кроссинговер. Это явление заключается в том, что хромосомные перестройки, произошедшие в одной паре хромосом, влияют на частоту перекреста в негомологичных хромосомах.
У кукурузы открыты гены, которые контролируют синапсис хромосом (асинаптический ген), спирализацию и слипание хромосом, а также установлен ген, препятствующий редукции хромосом в мейозе. Очевидно, что подобного типа гены могут влиять и на частоту обменов в гомологичных хромосомах. У ржи также обнаружен генетический контроль частоты образования хиазм.
Генотип влияет на частоту кроссинговера также и косвенно, через цитоплазму. У дрозофилы при реципрокном скрещивании установлено явление цитоплазматического эффекта на частоту перекреста.
Следует еще раз подчеркнуть, что частота кроссинговера является характерным показателем генотипа данной линии организмов. Намечается исключительно важная связь между частотой кроссинговера, характерной для генотипа, и другими генетическими явлениями, свойственными генотипу той же линии.
Влияние факторов внешней среды на перекрест хромосом
До сих пор мы рассматривали зависимость кроссинговера от генотипа и определяемого им физиологического состояния клеток и организма.
Такой тип перекреста называют спонтанным кроссинговером. Но частоту перекреста можно повысить, реже понизить влиянием на организм различных факторов внешней среды. Изменение частоты перекреста под влиянием факторов внешней среды называют индуцированным кроссинговером. Вероятно, сейчас нет достаточных основании разделять спонтанный и индуцированный кроссинговер, тем не менее изучение индуцированного кроссинговера может пролить свет на механизм перекреста хромосом и рекомбинацию генов.
На перекрест хромосом в профазе I мейоза влияют многие факторы внешней среды: высокая и низкая температура, ионизирующие излучения, инфракрасные лучи, дегидратация, отсутствие в растворах ионов кальция и магния и т. д.
Г. Плу, а затем К. Штерн и другие показали, что у дрозофилы низкие (9—13°) и высокие (30—32°) температуры увеличивают процент кроссинговера; в оптимальных температурных условиях развития обнаруживается наименьший процент перекреста.
Так, в исследованиях Плу учитывался кроссинговер между тремя рецессивными генами II группы сцепления, а именно
![]()
(с правой стороны вблизи гена pr находится центромера II хромосомы — отмечено стрелкой; цифры указывают локусы генов). Самки, гетерозиготные по данным генам, развитие которых проходило при разных температурах, были скрещены с самцами, гомозиготными по тем же рецессивным генам. В потомстве учитывались кроссоверные особи, возникшие вследствие перекреста Между генами b и pr и pr и c.

Зависимость частоты кроссинговера у дрозофилы от температуры
Частота перекреста значительно увеличивается при повышении и понижении температуры. Эти же материалы свидетельствуют о том, что действие температуры на перекрест в районе между генами b и pr, прилегающем к центромере, значительно больше (от 6,0 до 15,4% или 13,6%), чем между генами pr и c, где частота перекреста увеличивается всего на 6%.
Дальнейшими исследованиями было установлено, что районы хромосом вблизи центромеры более отзывчивы на внешние воздействия, чем удаленные от нее. Это явление связывают с более высокой реактивностью гетерохроматиновых районов вблизи центромеры.
Изучение действия рентгеновых лучей уже давно в работах Дж. Мэвора и К. Свенсона в 1924 г. и Г. Мёллера в 1925—1926 гг. показало, что ионизирующая радиация влияет на кроссинговер. Если температура влияет главным образом на кроссинговер только в тех клетках, которые проходят в момент воздействия на профазу I мейоза, то рентгеновые лучи влияют на кроссинговер, повышая его частоту в клетках, находящихся как в предмейотическом, так и в мейотическом состояниях. Частота кроссинговера зависит от дозы облучения. В таблице приведены данные С. Синха о частоте кроссинговера для генов вблизи центромерного участка аутосомы дрозофилы.
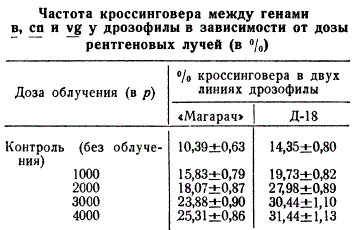
Однако указанная зависимость имеется только для определенной стадии развития половых клеток, соответствующей профазе I мейоза, и зависит также от генотипа линий и определенного района хромосомы (гетерохроматинового).
Исследование действия химических агентов также показало, что многие из них сходным с рентгеновыми лучами образом увеличивают частоту кроссинговера. К таким агентам относятся иприт (горчичный газ), формальдегид, органическая перекись, этиленимин и др.
Наиболее изученным из химических агентов по эффекту на кроссинговер является этилендиаминтетрауксусная кислота (сокращенно ЭДТА). Предполагается, что этот агент удаляет из хромосомы бивалентные ионы кальция и магния, которые, по-видимому, играют роль в поддержании структурной целостности хромосом. Предполагается, что удаление их ведет к нарушению непрерывности структуры хромосом, что и увеличивает частоту хроматидных разрывов, часть из которых может приводить к рекомбинации генов.
Механизм действия внешних факторов на кроссинговер все еще остается невыясненным.
Итак, резюмируя рассмотрение цитологических основ перекреста хромосом, можно сформулировать следующие положения.
1. Существуют два типа кроссинговера — митотический и мейотический.
2. Мейотический кроссинговер осуществляется в профазе I мейоза.
3. Началом осуществления перекреста является синапсис гомологичных хромосом на зиготенной стадии.
4. Перекрест происходит не между хромосомами, а между хроматидами, причем с равной вероятностью как между сестринскими, так и между несестринскими. Но перекрест между сестринскими хроматидами не обнаруживается.
5. В результате анализа механизма кроссинговера можно с определенностью сказать, что:
- кроссинговер осуществляется между генами, при этом ген оказывается как бы единицей кроссинговера;
- каждая рекомбинация происходит реципрокно как следствие обмена;
- каждый произошедший обмен в данном участке обнаруживается только в половине всех продуктов мейоза (т. е. в 50% гамет или спор);
- одинарные кроссинговеры могут захватить только две хроматиды; двойные и множественные перекресты могут захватывать две, три или все четыре хроматиды.
6. При изучении перекреста хромосом было выяснено, что первое и второе мейотические деления с генетической точки зрения являются одновременно и редукционными, и эквационными.
Итак, цитологический анализ перекреста хромосом позволил углубить наши знания по целому ряду генетических явлений, которые сначала были установлены только на основе скрещивания.
К сказанному следует добавить, что кроссинговер играет исключительно важную роль в процессе эволюции. Благодаря перекресту благоприятные гены могут комбинироваться в наиболее приспособительном для организма сочетании. В случае сцепления благоприятных генов с вредными, с помощью перекреста они могут разъединяться. Таким образом, кроссинговер обеспечивает своеобразную генетическую лабильность в процессе эволюции.