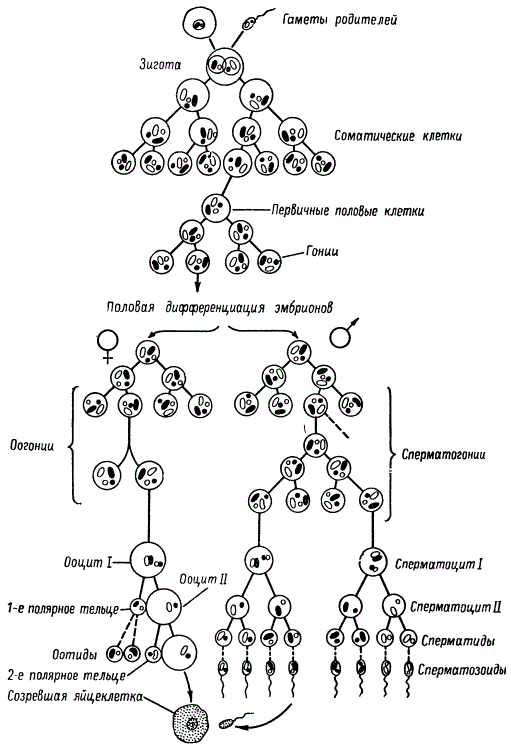
У животных половые клетки происходят из эмбриональных зародышевых клеток, из которых в результате многократного деления могут возникать как соматические, так и зародышевые клетки.
Последние путем ряда повторных делений дают первичные, или примордиальные, половые клетки, которые физиологически являются еще не дифференцированными. В ряде случаев они могут быть сходными с эмбриональными соматическими клетками. После периода покоя различной продолжительности примордиальные клетки превращаются в гониальные клетки — гонии. Вначале они сходны у особей обоих полов, но затем дифференцируются: у самцов — в первичные сперматогонии, а у самок — в первичные оогонии. При последующих митотических делениях они превращаются во вторичные сперматогонии и оогонии.
Первичные и вторичные сперматогонии и оогонии размножаются путем обычного митоза. В процессе ряда митотических делений при сохранении диплоидного набора хромосом они уменьшаются в размере и, наконец, перестают делиться. Клетки вступают в период роста, увеличиваются в размерах. На этой стадии развития незрелые мужские половые клетки с диплоидным набором хромосом называют сперматоцитами первого порядка (сперматоцит I), а женские — ооцитами первого порядка (ооцит I).
В формировании женских и мужских половых клеток существуют коренные различия.
Сперматогенез
Сперматоцит I вступает в период мейотического деления; у животных оно называется делением созревания.
Сравнительная схема развития мужских (сперматогенез) и женских (оогенез) половых клеток
В период роста сперматоцита I в его ядре проходят те профазные изменения, которые описаны выше при характеристике фаз мейоза. В результате первого деления созревания образуются сперматоциты второго порядка (сперматоцит II), которые имеют вдвое уменьшенное число хромосом, т. е. являются гаплоидным. При этом каждая из двух дочерних клеток (в норме) получает по одной гомологичной хромосоме каждой пары. После второго деления созревания — эквационного деления — из каждого сперматоцита II с гаплоидным набором хромосом образуется по две клетки также с гаплоидным набором хромосом. Эти клетки называют сперматидами. Итак, из одной диплоидной клетки — сперматоцита I, в результате двух мейотических делений образуются четыре гаплоидные сперматиды.
Процесс превращения сперматид в сперматозоиды называется спермиогенезом, или спермиогистогенезом. В нем участвуют все элементы ядра и цитоплазмы. Зрелый сперматозоид имеет головку, среднюю часть (шейку) и хвост. Ядро сперматиды преобразуется в головку сперматозоида. Цитоплазматические органоиды, структурно модифицируясь, превращаются в различные его органы, обеспечивающие движение сперматозоида и проникновение его в яйцеклетку. Центриоль входит в шейку и соединяет осевую ее часть с ядром головки. Различают проксимальную и дистальную центриоли. Митохондрии образуют комплекс, расположенный витками в районе шейки, а элементы аппарата Гольджи — акросому, или перфораторий. Цитоплазма в сперматозоиде представлена тонкой оболочкой.
Химический состав ядра сперматозоида сходен с хромосомным, но гистоны в нем замещены протаминами. Так как ядро сперматозоида содержит гаплоидное число хромосом, то количество ДНК здесь оказывается в два раза меньше, чем в ядре диплоидных клеток. Среднее количество ДНК на диплоидное ядро у разных видов животных в клетках разных тканей колеблется от 2,01⋅10-12 до 7,33⋅10-12 г.
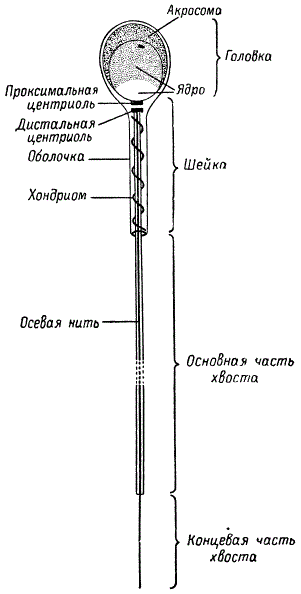
Схема строения сперматозоида млекопитающих
С помощью электронного микроскопа установлено, что ядро сперматозоида имеет кристаллическое строение, обусловленное параллельным расположением молекул дезоксирибону клеопротеида. Такое состояние может быть приспособительным для сохранения и переноса генов мужской особи, поскольку сперматозоид вне организма подвержен различным внешним воздействиям.
Действительно, исследования последних лет показали, что незрелые половые клетки и сперматиды более чувствительны к воздействию внешних факторов, в том числе и к действию ионизирующей радиации, чем зрелые сперматозоиды.
По своей морфологии сперматозоиды чрезвычайно разнообразны и характерны для каждого вида. Например, недавно Дж. Тейлор авторадиографическим методом установил, что в сильно вытянутой головке сперматозоида кузнечика Romalia microptera хромосомы располагаются цепочкой одна за другой, образуя как бы одну вытянутую хромосому. Морфологические отличия сперматозоидов разных видов способствуют их половой изоляции, препятствуя смешению видов.
Оогенез
В принципе путь развития женских половых клеток сходен с таковым у мужских.
Во-первых, стадия роста ооцитов первого порядка (ооцит I) более продолжительна, чем сперматоцитов первого порядка, так как в этот период в ооците — будущей яйцеклетке происходит накопление питательных веществ.
Во-вторых, из каждого ооцита I после двух мейотических делений хотя и образуются четыре оотиды, но только одна из них (яйцеклетка) способна к дальнейшему развитию и слиянию со сперматозоидом. Другие три оотиды оказываются абортивными, они даже не обособляются в полноценные самостоятельные клетки. Эти три оотиды, также с гаплоидным набором хромосом, но без достаточного запаса цитоплазмы, не способны обеспечить развитие зиготы. Они называются редукционными, или полярными, тельцами и образуются следующим образом. После первого деления созревания ядро одного тельца (первое полярное тельце) отходит к периферии, где претерпевает второе деление, образуя две оотиды.
В результате второго деления созревания яйцеклетки возникает второе полярное тельце, т. е. третья оотида. В редких случаях наблюдается, что одна из трех оотид соединяется с ядром одного из проникших в нее сперматозоидов и одновременно с оплодотворенной яйцеклеткой дает начало организму с наследственно разными тканями (химеры). Такое явление наблюдается, например, у тутового шелкопряда. Следовательно, мейотические деления в ооцитах, в отличие от таковых в сперматоцитах, не сопровождаются равным делением тела клетки. В типичном случае из четырех клеток, возникающих в результате двух мейотических делений, только одна превращается в яйцеклетку.
Изучение истории развития половых клеток животных показало, что они происходят из недифференцированных соматических клеток. Процесс обособления зачатковых клеток в онтогенезе, из которых впоследствии развиваются половые железы и половые клетки, называется зачатковым путем. У одних животных он закладывается в процессе формирования эмбриона (млекопитающие, человек), у других — на еще более ранних стадиях. Так, Т. Бовери показал, что у лошадиной аскариды (Ascaris megalocephala) уже при первом дроблении оплодотворенного яйца бластомеры различаются по состоянию хромосом: один из двух бластомеров при дальнейшем дроблении яйца и развитии зародыша дает группу клеток, из которой образуется половая железа, а затем и половые клетки, в то время как из другого бластомера развивается тело (сома) организма.
У насекомых, ракообразных и червей дифференцирование клеток в онтогенезе на половые и соматические происходит также очень рано. Эти явления указывают на раннюю специализацию половых клеток, предназначенных для передачи наследственных свойств следующему поколению.