Генетика, владея возможностью произвольного создания соответствующих генотипов, предложила свои подходы к изучению наследственной детерминации онтогенеза.
К ним относятся исследования:
- молекулярного характера действия генов в цепи, биосинтеза ДНК — РНК — белок;
- мутантных признаков, начала их дифференцировки, времени и характера проявления;
- взаимодействия мутантных аллелей и генов дикого типа в генотипе;
- мутаций и рекомбинаций соматических клеток в процессе дифференциации тканей и морфогенеза;
- возникновения фенокопий, т. е. фенотипических изменений, сходных по проявлению с мутациями;
- развития отдаленных гибридов, у которых сочетаются признаки разных видов;
- проявления признаков у однояйцевых близнецов;
- влияния сходства и различия генотипов на приживление тканей при гетеро- и гомопластических пересадках тканей;
- иммунологических реакций у генотипически различающихся организмов;
- биосинтеза на основе биохимических мутантов;
- влияния генома и плазмона на развитие признака;
- проявления генетической нормы реакции в различных условиях внешней среды, и др.
Основным и исходным моментом генетических исследований Онтогенеза является возможность дискретного анализа действия гена на формирование признака: один ген — один признак. Современное представление этого положения можно записать: один ген (ДНК) — РНК — белок — признак.
Главной проблемой генетики в изучении наследственных основ индивидуального развития теперь является установление промежуточных звеньев в цепи ген — признак. Однако для того чтобы понять механизм действия гена.-взаимосвязь гена и среды, необходимо помнить, что сам ген непосредственно не производит специфического белка, следовательно, и не определяет конкретного признака. Развитие признака как детали многоклеточного организма определяется всей системой генотипа в процессе многоступенчатых и взаимосвязанных актов. Именно поэтому действие гена может быть изучено лишь в системе онтогенеза, касается ли это многоклеточного или одноклеточного организма.
Изучение генетики онтогенеза ведется на трех уровнях: молекулярном, клеточном и организменном. При этом основными методами исследования остаются генетические, благодаря которым можно получать различные комбинации генотипов и геномов.
Согласно принятому в эмбриологии определению, в понятие онтогенеза включают процесс развития, начинающийся с акта оплодотворения или активации развития яйца и заканчивающийся смертью организма. Процесс онтогенеза животного состоит из качественно различных периодов — эмбрионального и постэмбрионального; в последнем можно выделить этапы роста и полового созревания. Аналогично этому, несколько этапов качественных изменений претерпевают и растения.
В ходе развития животных и растений, хотя и своеобразно, осуществляется ряд сходных основных процессов: 1) рост, 2) дифференцировка тканей, 3) морфогенез, т. е. развитие органов и признаков. При этом непрерывно происходит биохимическая, физиологическая (функциональная) и морфологическая дифференцировка тканей и органов. В процессе развития создается ряд механизмов, обеспечивающих приспособление организма к окружающей среде. Теперь становится очевидным, что в онтогенезе происходит и генетическая дифференцировка соматических тканей, так как установлено, что разные ткани различаются по состоянию хромосом, набору хромосом и функционирующих генов. Поэтому разные ткани имеют различный биохимический состав соответственно функции ткани и органа.
Для изучения генетики онтогенеза совершенно необходимо учитывать общие закономерности онтогенеза, которые установлены эмбриологами. К таким закономерностям относятся следующие:
1. Прохождение в эмбриогенезе фаз развития, характерных для предков, что указывает на генетическую предопределенность общего плана развития данного класса животных и на общность происхождения (биогенетический закон);
2. Неравномерность прохождения процессов роста и дифференцировки: в период роста преобладают процессы, связанные с митотической активностью клеток, в период дифференцировки усиленно функционируют гены, определяющие синтез специфических белков, необходимых для формообразовательных процессов и интеграции специализированных клеток;
3. Необратимость и обратимость дифференцировки соматических клеток и тканей. Необратимость дифференцировки имеет место в тех случаях, когда генетическая специализация ядер и цитоплазмы в клетках уже произошла и они утратили способность воссоздавать целый организм.
Однако широко известны случаи, когда группа клеток или отдельная клетка уже дифференцированной ткани сохраняют способность путем дезинтеграции и последующей новой дифференцировки восстановить целый организм или часть его. Это служит доказательством того, что ядра и цитоплазма этих клеток генетически не дифференцировались. Такую обратимую дифференцировку обычно называют тотипотентностью. Но нам кажется, что более правильно это явление называть соматическим эмбриогенезом, как это было предложено в 1959 г. Б. П. Токиным.
4. Индукционность отношений между тканями в период эмбриогенеза. Изменение морфогенеза путем пересадки генетически детерминированного участка, например губы бластопора, на новый участок эктодермы другого зародыша индуцирует (организует) развитие здесь того органа и сопутствующего ему комплекса, который обычно им детерминируется.
5. Параллелизм рядов гистологической структуры и функции тканей (нервной, мышечной, эпителиальной и соединительной) в разных филогенетических рядах. Наличие этой закономерности, установленной А. А. Заварзиным, отражает древность и общность генетической детерминации клеток в весьма давно разошедшихся классах, например у млекопитающих, относящихся к вторичноротым, и у насекомых, относящихся к первичноротым.
Все эти и другие более частные закономерности онтогенеза являются предметом изучения эмбриологии. Одни исследователи считают, что процесс дифференцировки целиком преформирован генами, другие отрицают значение генов в этом процессе, признавая ведущую роль за цитоплазмой и факторами внешней среды. Ни одна из этих крайних точек зрения не является правильной. Начиная с первого деления зиготы, все клетки организма содержат одни и те же наборы геномов, и тем не менее в эмбриогенезе происходит закладка неодинаковых тканей и органов с различными функциями и не одновременно, а в строгой последовательности.
Из эмбриологии известно, что первичная морфологическая дифференцировка обусловлена структурой цитоплазмы и поверхностным, или кортикальным, слоем яйца (кортексом).
Были приведены факты, свидетельствующие о том, что безъядерные яйца амфибий и некоторых беспозвоночных после их активации сохраняют способность развиваться до стадии бластулы. Подобные опыты служили иногда поводом для признания того, что первые стадии развития эмбриона не зависят от действия генов и обусловлены строением цитоплазмы яйца.
На этом основании некоторые эмбриологи считают, что цитоплазма и кортекс несут в себе самостоятельную закодированную информацию дифференцировки, которая затем декодируется. При этом допускается, что заключенные в них формообразовательные аппараты способны сохранять программу дифференцировки видовых признаков. На долю ядерных генов они отводят определение признаков индивидуального характера.
Кортекс имеет функциональную зональность: анимальную, из которой образуется эктодерма, зону «серого серпа», где закладывается мезодерма и откуда начинается гаструляция, и вегетативную, образующую эндодерму. Кортикальный слой яйца определяет полярность и дорзовентральную ориентацию яйца и зародыша. Яйцо оказывается дифференцированным еще до оплодотворения. После оплодотворения выявляется еще более тонкая дифференциация яйца, детерминирующая ход развития зародыша на ранних стадиях эмбриогенеза.
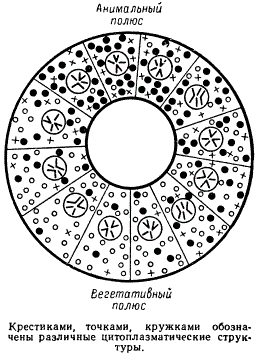
Схема, иллюстрирующая неравномерность распределения цитоплазматических структур в процессе дробления яйца
Бластомеры вследствие митотического деления содержат одинаковые геномы, но размеры участков кортекса и цитоплазмы в них неодинаковы. В процессе дробления яйца разные бластомеры имеют различно преформированную цитоплазму, которая может служить регулятором считывания разных генов в различных бластомерах и таким образом влиять на ход дифференцировки.
Это предположение достаточно обосновано и может служить хорошей гипотезой взаимосвязи ядра и цитоплазмы в процессе дифференцировки. Но при этом нельзя не обратить внимание на то, что преформированность цитоплазмы и кортикального слоя яйца является результатом деятельности генотипа материнского организма.
Мы уже приводили целый ряд примеров генетического детерминирования цитоплазмы, влияющей на развитие признаков потомства: левая и правая закрученность завитка раковины моллюска, цитоплазматическая мужская стерильность кукурузы, развитие отдаленных гибридов при несовместимой с ядром цитоплазме (на примере дрозофилы) и т. д. Свидетельством того, что генотип матери способен самостоятельно обеспечить нормальное воспроизведение потомства, служит также тот факт, что у животных и растений широко распространен партеногенез.
Следует иметь в виду, что геномы матери, формирующие структуру яйца, не совпадают с геномами потомства. В формировании яйца принимает участие весь набор генов диплоидного материнского организма. После мейоза же в яйце сохраняются гены лишь гаплоидного набора хромосом. Однако в цитоплазме и кортикальном слое остаются все материнские генные продукты и сформированные структуры. Именно они обеспечивают начальные фазы развития яйца, представляя собой информацию, заранее заготовленную материнским организмом. И поскольку в диплоидном материнском организме присутствуют материнский и отцовский наборы хромосом и генов данного вида, то этим путем и обеспечивается реализация плана онтогенеза.
Таким образом, преемственность детерминации онтогенеза может осуществляться за счет действия не только наличных генов зиготы, но и генных продуктов материнского генотипа. В этом случае запоминание того, что должно происходить в начале онтогенеза, осуществляется не генами, а генными продуктами и структурами яйца.
Функционирование генных продуктов в клетке в отсутствие ядра имеет место не только в яйце. Примером этому служат безъядерные эритроциты млекопитающих. Утрата ядра красных кровяных телец происходит при формировании ретикулоцитов, но функционирование его продуктов сохраняется и в эритроцитах.
Механизм генетической памяти в онтогенезе не обязательно связан с необходимостью синтеза и-РНК в каждый данный момент в каждом данном состоянии клетки. Предварительное избыточное накопление и-РНК может обеспечивать какое-то время синтез специфических белков в отсутствие ДНК. Следовательно, генетическая дифференцировка яйцеклетки может быть следствием предшествующего функционирования генов матери и сохранения информационной РНК, способной служить матрицей синтеза белков в ходе дифференцировки и развития зародыша. Кроме того, следует иметь в виду, что органоиды клетки имеют собственные нуклеиновые кислоты — ДНК и РНК, которые, быть может, также способны кодировать и передавать информацию, как и ядерные нуклеиновые кислоты.
Образованием готовой к оплодотворению яйцеклетки заканчивается первый этап онтогенеза.
С оплодотворением начинается новый этап онтогенеза, в котором скажется действие генов отцовского организма. Но поскольку гены матери, определяющие структуру яйца, реализуются в самом начале онтогенеза, их плейотропный эффект на последующих этапах дифференцировки проявляется сильнее, чем эффект генов отца. Ведь совершенно очевидно, что если группа генов определяет закладку зародышевых листков (эктодермы или эндодермы), то влияние этих генов скажется в более широких и глубоких последствиях на ходе развития по сравнению с теми генами, которые вступают в действие позднее. С генетической точки зрения можно сформулировать общий принцип: чем раньше проявляется действие генов в онтогенезе, тем значительнее их плейотропный эффект. Процесс дифференцировки зародыша, как предполагают, определяется механизмом считывания разных генов по мере специализации клеток различных тканей.
Если гены действительно контролируют весь план развития и соответственно все признаки и реакции организма, то возникает ряд вопросов:
- на разных этапах онтогенеза действуют одновременно все гены или разные?
- чем определяется вступление их в действие?
- каким образом осуществляется специфическое действие генов?
Для изучения генетической детерминации онтогенеза применяют различные методы: метод трансплантаций, цитогенетические, биохимические, иммунологические, физиологические и другие методы.
Так, при помощи трансплантации ядер исследуется эквипотенциальность клеток. Метод состоит в следующем: неоплодотворенные яйца, например, лягушки активируют уколом и удаляют из них ядра. Затем с помощью микропипетки в каждое из таких яиц пересаживают по ядру из клеток зародышей, находящихся на различных стадиях развития (бластулы, гаструлы или на еще более поздних стадиях). В том случае, если ядро клетки-донора уже претерпело какую-то дифференциацию в процессе дробления, то после его пересадки яйцо-реципиент не даст нормального зародыша. Если ядро донора в процессе клеточных делений еще не было дифференцировано и сохранило исходную потенцию — способность обеспечивать полное развитие (тотипотентность), то яйцеклетка-реципиент будет нормально дробиться и даст головастика. Подобными опытами было показано, что если ядро берется от клетки-донора со стадии бластулы или ранней гаструлы, то из яйца-реципиента развивается нормальный головастик. Следовательно, ядра клеток на ранних стадиях развития еще не дифференцированы и эквипотенциальны ядру зиготы в его исходном состоянии. Из яйца с ядром, пересаженным из клеток поздней гаструлы, зародыш не развивается: развитие блокируется на стадии бластулы или гаструлы. В данном случае ядро клетки-донора в стадии поздней гаструлы оказывается уже дифференцированным, т. е. претерпевшим необратимые изменения в ходе развития.
Следует, однако, заметить, что ядра клеток некоторых тканей в процессе дифференцировки зародыша довольно долго сохраняют тотипотентность и при пересадке их в безъядерное яйцо обеспечивают его нормальное развитие.
Цитогенетическими методами изучают состояние и гетероциклические изменения отдельных районов хромосом в клетках различных тканей.
В процессе дифференцировки и функционирования соматических клеток хромосомы претерпевают значительные изменения, связанные со спецификой обмена веществ. Принято считать, что гены функционируют только в интерфазном состоянии ядра клетки; в метафазе они не функционируют, т. е. не выделяют в цитоплазму генных продуктов. Предполагается, что функционирование гена происходит в состоянии деспирализации хромосом.
В клетках некоторых тканей растений (перидерма, клетки тапетума и др.) и животных (мальпигиевы трубки, кишечный эпителий и др.) часто наблюдается явление эндомитоза, в результате которого клетки оказываются полиплоидными, что, очевидно, связано с особенностями дифференцировки этих тканей. В клетках дифференцированных тканей некоторых видов животных и растений наблюдаются явления амитоза, политении, полиплоидизации и соматической редукции хромосом. Например, в клетках эпителия кишечника комара (Culex pipiens) наблюдается сначала умножение числа хромосом (полиплоидизация), а затем — конъюгация гомологичных хромосом и их расхождение в митозе без продольного расщепления. Таким образом, здесь происходит редукция числа хромосом как закономерный процесс в ходе дифференцировки кишечного эпителия. Политенное состояние хромосом, как мы знаем, характерно для ядер слюнных желез личинок двукрылых. Политенные хромосомы обнаруживаются у них также в ядрах клеток кишечника, мальпигиевых трубок.
Все эти примеры указывают на несомненную связь состояния ядра и хромосом с морфофизиологической дифференцировкой соматических клеток организма. Правда, остается неясным главный вопрос, с какими конкретными функциями клетки связаны те или иные изменения состояния ядра и хромосом, но решение этих вопросов — дело ближайшего будущего.
В последние годы цитогенетикам удалось установить очень интересное явление локального изменения состояния отдельных участков хромосомы (гетероцикличность хромосом) в зависимости от стадии развития организма. Прослеживая состояние одних и тех же дисков гигантских хромосом в клетках слюнных желез личинок двукрылых (хирономуса, дрозофилы), исследователи обнаружили, что в определенных местах хромосомы на определенной стадии развития на месте некоторых дисков появляются вздутия (по английски — puffings). Хромосомные нити в этих участках деспирализованы, т. е. приобретают состояние, характерное для интерфазного функционирующего ядра.
Это состояние непостоянно и обратимо. Показано, что характер вздутий меняется в разные периоды жизни личинки в различных участках железы, причем каждый диск имеет свое «расписание» образования вздутий. Замечено, что возникновение и исчезновение их связано с динамикой нуклеиновых кислот. Большой интерес представляют работы, где показано, что в хромосомах при пересадке ядер из клеток слюнных желез предкуколки в клетки зародыша, находящегося на более ранних этапах развития, изменяется место появления вздутий.
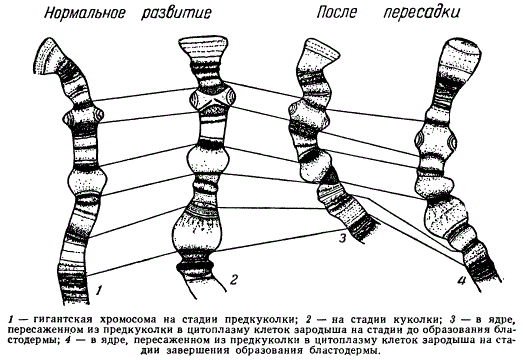
Изменение места появления вздутий на гигантской хромосоме у Drosophila при нормальном развитии и после пересадки
Конечно, эти локальные изменения хромосом еще очень мало говорят о характере действия гена, но такого рода исследования расширяют наши подходы к изучению роли хромосом и генов в функционировании клетки.
Итак, генетическое и цитогенетическое изучение морфофизиологической дифференцировки тканей с привлечением новых методов исследования дает основание сделать следующее заключение. В процессе онтогенеза клетки соматических тканей претерпевают дифференцировку. При этом ядра соматических клеток уже на ранних стадиях эмбриогенеза теряют свою исходную тотипотентность, а хромосомы локально изменяют состояние спирализации.