У плодовых и ягодных растений отдаленная гибридизация — один из важнейших факторов видообразования как в природе, так и в культуре. Об этом писали Н. И. Вавилов, И. В. Мичурин, Л. Бербанк и другие ученые. Процесс особенно активно идет, когда молодые виды осваивают новые территории и в процессе миграции сталкиваются с родственными видами. Отдаленная гибридизация имеет важное значение при становлении плодовых культур, когда в ходе одомашивания и интродукции происходит совместное произрастание видов, ареалы которых в природе не совмещаются.
Гибридогенные (возникшие в результате гибридизации) виды плодовых и ягодных растений молоды в эволюционном плане, они активно приспосабливаются к изменениям внешней среды в процессе расширения своих ареалов. Возникают сорта с высокой степенью адаптивности.
Пути возникновения гибридогенных видов у плодовых и ягодных растений неодинаковы. При наложении ареалов дикорастущих видов отдаленная гибридизация между ними приводит не только к интрогрессии отдельных генов или блоков генов от одних видов к другим, не меняя их видовой или родовой тип, но и к возникновению гибридогенных видов со своими оригинальными комплексами признаков.
В результате спонтанной гибридизации в природе возникли виды миндаля: миндаль колючейший — A. spinosissima Bge. (миндаль метельчатый — A. scoparia x микровишня мелкоплодная — M. microcarpa), миндаль бухарский — A. bucharica (миндаль обыкновенный — A. communis x микровишня мелкоплодная), миндаль Фенцля — A. fenzliana (миндаль обыкновенный — A. communis х алыча — Р. cerasifera), миндаль низкий — А. nana (миндаль метельчатый — A. scoparia х микровишня седая — М. incana), миндаль Петунникова — A. petunnicovi (миндаль низкий — А. nana x миндаль колючейший — А. spinosissima Bge.). Они возникли в местах наложения ареалов засухоустойчивых среднеазиатских видов с переднеазиатскими мезофитными (или ксеромезофитными) видами. Это объясняет исключительно высокую адаптивность гибридогенных видов миндаля к абиотическим стрессам, что помогло им занять новые для рода Amygdalus территории, где их предковые виды не способны произрастать.
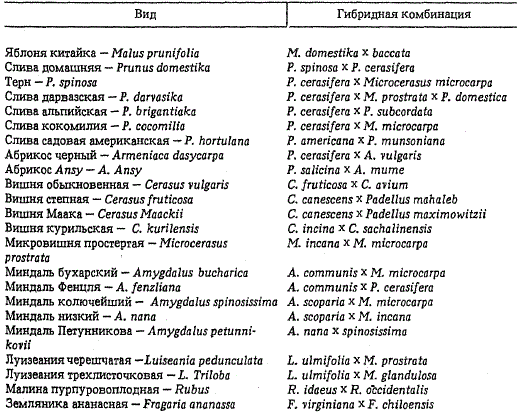
Происхождение гибридогенных видов плодовых и ягодных растений
Во всех случаях гибридогенные виды занимают экологическую нишу, сходную с родоначальным видом того же рода. Миндаль бухарский и Фенцля — среднегорья, как и миндаль обыкновенный; миндаль колючейший и низкий — низкогорья до границы со степью, как и миндаль метельчатый.
Гибридогенные дикорастущие виды плодовых и ягодных растений выделены во всех центрах происхождения. В Восточноазиатском генцентре гибридное происхождение установлено для таких видов, как абрикос маньчжурский — A. mandshurica (абрикос китайский — А. mume х микровишня железистая — М. glandulosa), вишня курильская — C. kurilensis (вишня сахалинская — С. sachalinensis х вишня — С. incisa), вишня Маака — С. maacki (вишня — С. canescens х паделлус Максимовича — Р. maximovitzi), луизеания трехлисточковая — L. triloba (луизеания вязолистная — L. ulmifolia х микровишня железистая — М. glandulosa).
В Средней Азии при совпадении ареалов аборигенных видов и мигрантов из Восточноазиатского и Переднеазиатского генцентров возникли такие гибридные виды косточковых растений, как микровишня простертая — М. prostrata (микровишня мелкоплодная — М. microcarpa х микровишня седая — М. incana), слива дарвазская — Р. darvasika (слива домашняя — Р. domestica х микровишня простертая — М. prostrata), луизеания черешчатая — Р. pedunculata (вишня простертая — М. prostrata х луизеания вязолистная — L. ulmifolia), а также миндаль бухарский и колючейший.
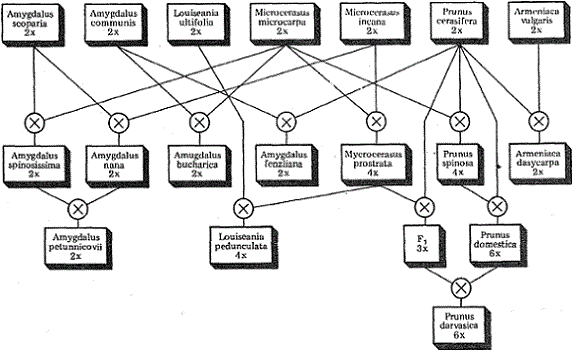
Отдаленная гибридизация и видообразование у косточковых растений на стыке среднеазиатского, переднеазиатского, восточноазиатского и европейского генетических центров
В Переднеазиатском генцентре возникновение гибридогенных видов связано с деятельностью человека. Поскольку эти виды возникли в культуре, их называют культигенными. В дикорастущем виде они встречаются только на месте заброшенных садов, в одичавшем состоянии.
Пути возникновения таких гибридогенных видов различны. Некоторые из них возникли в результате интрогрессивной гибридизации с несколькими родственными видами. Биологический и морфологический тип при этом остается близким к первоначальному виду, на основе которого возник вид культигенный. Первоначальный, или предковый, вид в этом случае получил название базисного. Признаки новых культигенных сортов изменились по сравнению с базисным видом (реципиентом) настолько, что имеются все основания считать его самостоятельным культигенным видом. Многие культигенные виды отличаются большим полиморфизмом, у разных сортотипов преобладают признаки различных видов-доноров, поэтому они получили название сборных. К числу таких сборных культигенных видов относятся наши важнейшие плодовые растения.
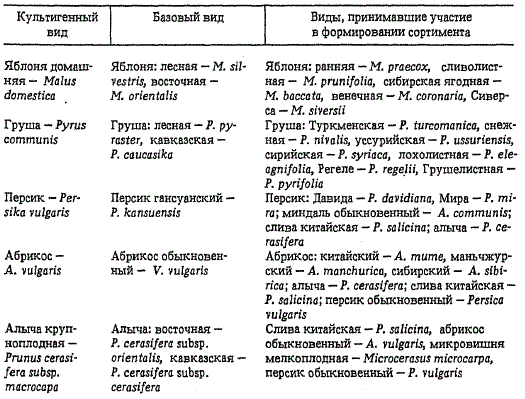
Происхождение плодовых культур
При гибридизации между видами плодовых растений в условиях культуры возникают стабильные культигенные виды, получившие широкое распространение: земляника ананасная — Fragaria ananassa Dubh. (земляника виргинская — F. virginiana Mill, х земляника чилийская — F. chiloensis Duch.), малина пурпуровоплодная — Rubus (малина красная — R. idaeus L. х малина черная — R. occidentalism), вишня обыкновенная — Cerasus vulgaris Mill. (вишня — С. avium х вишня степная — С. fruticosa), слива домашняя — Р. domestica (алыча — Р. cerasifera х терн — Р. spinosa), абрикос Ансу — A. Ansu (Maxsim) Kosh. (абрикос китайский — А. mume Sieb. Х слива китайская — Р. salicina), абрикос черный — A. dasycarpa Pets. (алыча — Р. cerasifera х абрикос обыкновенный — A. vulgaris), яблоня сливолистная — M. prunifolia (яблоня домашняя — Р. domestica х сибирка — М. baccata).
Все эти виды хорошо сохраняют родовой морфологический и биологический тип материнского вида, но признаки другого компонента гибридизации выражены достаточно четко. Они константны, очень пластичны. Некоторые из них значительно превосходят свои родительские виды по хозяйственным и биологическим показателям, что сделало их ведущими видами культуры (земляника ананасная, вишня обыкновенная, слива домашняя). Эти виды перспективны в качестве улучшителей при интрогрессивной гибридизации.