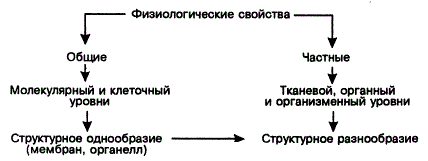
Физиологические свойства как наследственно дискретные особенности жизнедеятельности растений чрезвычайно разнообразны и могут быть разделены на общие и частные.
К общим относятся способность к фотосинтезу и аэробному дыханию, зависимость продуктивности и размножения растений от изменения среды, наличие зеленого пигмента и т. д. Частные свойства проявляются в ограничении транспирации (суккуленты) и высоком осмотическом давлении (галофиты), в предпочтительном поглощении нитратного или аммонийного азота разными растениями, в специфике фотопериодической реакции, специфике покоя и т. д.
Общие и частные (специфические) физиологические свойства могут быть разложены на отдельные составляющие. Так, путь метаболизма углерода у растений состоит из множества реакций, каждая из которых важна для синтеза органического вещества и поддержания жизнедеятельности. Транспирация растений складывается из особенностей метаболизма устьичных клеток, состояния оводненности клеток листа, выделительной деятельности клеток и т. д.
Физиологические свойства реализуются на молекулярном, клеточном, тканевом и органном уровнях, а также на уровне целого растения. С переходом от молекулярного к организменному уровню, особенно у многоклеточных, физиологические свойства приобретают более частный характер. Так, молекулярные процессы жизнедеятельности всех растений носят более. общий характер, чем процессы на органном и организменном уровнях. Это и есть одно из проявлений физиологической дифференциации видов. Напомним, что общие молекулярные механизмы фотосинтеза и поглощения веществ могут иметь специфические особенности на тканевом, органном и организменном уровнях. Так, клетки не всех тканей и органов ассимилируют CO2 подобно клеткам палисадной ткани листа. Точно так же не все растения, способные к фотосинтезу, осуществляют его в листьях.
Физиологические свойства — интегральный результат молекулярных и морфологических изменений. Первичной основой функциональной специфики являются молекулярные процессы, осуществляющиеся на мембранах и в органеллах, а их конечная реализация зависит от сложной и разнообразной структуры (тканей, органов, систем органов). Поэтому структурная дифференциация оказывается предпосылкой для дальнейшей функциональной эволюции. Это еще одно свидетельство неправомерности ограничения функциональной эволюции молекулярными процессами.
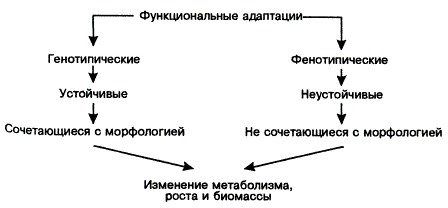
Все это имеет отношение и к классификации функциональных адаптаций растений. Вопрос о классификации адаптаций — один из нерешенных в эволюционном учении. Наиболее удачной признана пока классификация адаптаций по масштабу, происхождению, принадлежности к разным средам и т. д. Функциональные адаптации растений можно разделить на генетические (устойчивые) и фенотипические (неустойчивые).
К устойчивым, как правило, относят адаптации, сочетающиеся с наследственными изменениями в морфологии. Например, наличие обкладки пучка сочетается с изменением пути фиксации CO2 (C4-путь фотосинтеза), а устойчивость к засухе определяется комплексом признаков. Так, устойчивость к обезвоживанию может быть усилена изменением мембранной активности, осмотических сил клеток и корня, индуцированием развития световых листьев, часть которых является модификациями, но тем не менее они важны для выживания растений. Нередко их обозначают термином акклимация, чтобы отграничить от наследственно фиксированных адаптаций (N. Turner, Р. Kramer, 1980).
Функциональные адаптации не всегда сопровождаются видимыми морфологическими перестройками, например суточная и сезонная динамика фотосинтеза и транспирации, Изменение интенсивности дыхания при временных перепадах температуры и т. п. Многие адаптации функционального характера морфологически выражены, отражаются на росте растений и органов, соотношении подземной и надземной биомассы, на числе клеток и сосудов.
По специфике и рангу функциональные адаптации могут быть разделены на группы:
- проявляющиеся стабильно и общие для всех растений (реакции циклов Кальвина и Кребса, клеточные основы и полярность роста и т. д.);
- развивающиеся в строго определенных условиях и чередующиеся в онтогенезе (суточные и сезонные ритмы, фотопериодическая реакция, покой и т. д.);
- необходимые для переживания стрессовых условий, отражающиеся на структуре (последствия засухи, засоления и т. д);
- реализующиеся в течение короткого времени для обеспечения одного из этапов онтогенеза (функция нектарников, зародышевых корней и семядолей, реакция на опыление и т. д);
- ненаследственные и обратимые свойства-модификации (временные изменения обменных процессов, морфологическая пластичность организма и т. д.), имеющие также значение для выживания организма.
Признаки 1—4-й групп наследственно закреплены и эволюционно стабилизированы.
Возможны и другие принципы классификации функциональных адаптаций, например по факторам воздействия и результатам. Приведенная схема, несмотря на ее упрощенность, дает возможность понять происхождение и значение адаптаций, что важно для последующего изложения материала.
Не совсем ясны параметры приспособленности, хотя чаще всего обращают внимание на выживаемость, репродуктивный успех, соответствие строения органа его функциям и т. п. Эти параметры, пригодные для организма и особенно вида, применительно к клетке и ее химизму не всегда себя оправдывают. Приходится считаться и с онтогенетическими адаптациями типа модификаций, довольно распространенными у растений. В частности, на примере бактерий и лиственницы показано, что линии, культивируемые в разных условиях, отличаются по температурному оптимуму фотосинтеза, что объясняется «закалкой» элементов функциональной адаптации (В. Я. Александров, 1985).
С аналогичным явлением мы сталкиваемся и при оценке температурной зависимости темнового дыхания листьев пшеницы, выращенных при различной температуре среды.