«Деревья, соединяясь в насаждение, — писал Г. Ф. Морозов, — вносят еще один новый элемент, отсутствующий у деревьев, изолированно друг от друга растущих, хотя бы число их было несметное. К двум другим функциям листовых органов (дыхание и ассимиляция) насаждение, по верному замечанию Бюсгена, присоединяет еще третью: листья после своего отпада дают в лесу так называемую лесную подстилку, иногда еще называемую лесным войлоком, а также мертвым покровом».
Далее Г. Ф. Морозов писал, что лесная подстилка «играет доминирующую роль в вопросе о влиянии леса на почву; весь химизм лесных почв, насколько он обусловлен лесом, весь подзолообразовательный процесс коренится, главным образом, в свойствах этой подстилки и условиях ее перегнивания; и если раньше старые лесоводы придавали ей громадное лесоводственное значение, исходя из односторонних представлений о ее положительной роли и как источника непосредственного питания (до Либиха) и как мертвого покрова, защищающего почву от испарения влаги, то теперь, после всестороннего освещения ее роли, когда ясно показано классическими исследованиями Мюллера, а затем Эмейса, Раманна и польза и вред ее, е зависимости от типа подстилки — значение ее в жизни леса еще более возросло».
Между строением лесных подстилок и характером перегноя лесных почв существует тесная зависимость. Каждому типу лесной подстилки соответствует определенный характер строения перегнойных горизонтов почвы, отличный по содержанию и качественному составу перегноя. Это послужило основанием для распространения наименований типов лесных подстилок на соответствующие им типы почвенного перегноя.
Первое встреченное в литературе указание на различный характер перегноя, образуемого под воздействием лесов разного породного состава, принадлежит профессору натуральной истории Московского университета М. И. Афонину. В 1771 г. М. И. Афонин на собрании Московского университета произнес торжественное слово, в котором давал следующее деление перегноя или, по его определению, чернозема:
- лесной, на смолу похожий;
- березовый, который сух и холоден;
- можжевелевый и сосновый, который натурою весьма горяч;
- еловый, который также жирен, но несколько холоднее соснового.
В 1826 г. Шпренгель писал о типах перегноя, образуемого при различных условиях разложения растительных остатков. В 1875 г. Эмейс выделил три главных типа перегноя, дав им следующую характеристику: в одном типе перегноя органическое вещество более или менее тесно смешано с минеральной частью поверхностного слоя почвы (вследствие чего можно предполагать возможность развития процесса нитрификации), в двух других типах органические остатки накапливаются главным образом на поверхности. Эти исследования подготовили основу для дальнейшей разработки классификации лесных подстилок и перегноя лесных почв.
Датским лесоводом П. Мюллером в 1887 г. была опубликована классификация лесных подстилок, оказавшая исключительно большое влияние на их последующее изучение, которая сохранила значение по настоящее время.
На основании изучения буковых, дубовых и еловых лесов Дании П. А. Мюллер установил два главных типа перегноя лесных почв: мягкий перегной, или муль, и грубый перегной, или мор.
В первом издании работы П. Мюллера, вышедшем на датском языке, был употреблен термин «mor», который в последующем немецком издании был переведен как «torf», в дальнейшем его переводили термином «Rohhumus», — сырой или грубый перегной.
Образование этих двух типов перегноя П. Мюллер связывал с характером растительности напочвенного покрова и с деятельностью червей, насекомых и других животных, населяющих почву.
Типичный муль, согласно исследованиям П. Мюллера, образуется в буковых лесах, где сплошной моховой покров отсутствует и лишь изредка встречаются разрозненные подушки мхов. Для напочвенного покрова этих лесов особенно характерен ясменник пахучий (Asperula odorata), затем перелеска многолетняя (Mercurialis perennis), бор развесистый (Milium effusum), перловник (Melicia uniflora), звездчатка дубравная (Stellaria holostea), отчасти кислица (Oxalis acetosella), ветреница дубравная (Anemone nemorosa). Почва покрыта рыхлым небольшой мощности слоем листьев, мелких сучьев, почечных чешуек, цветочных сережек и т. д. Граница между этим верхним слоем и лежащим под ним перегнойным горизонтом очень резкая. Перегнойный горизонт имеет черную или серовато-бурую окраску, зернистую структуру и очень рыхлое сложение. Верхнюю часть этого горизонта П. Мюллер называет «муль».
Микроскопический и механический анализ показывает, что в’ верхней части перегнойного горизонта различные органические остатки растительного и животного происхождения еще сохраняют ясно заметную структуру, в нижней части горизонта однородная темно-серая или буроватая масса перегноя равномерно обволакивает минеральные частицы почвы. Содержание перегноя 5—10%, постепенно снижающееся с глубиной. Водорастворимых органических веществ Мюллером не было обнаружено, и кислая реакция почвенного раствора в некоторых почвах с этим типом перегноя обусловлена влиянием углекислоты.
П. Мюллер обращает внимание на сильное развитие в рассматриваемом типе перегноя грибного мицелия. Исследованиями Рострупа было обнаружено до 47 видов крупных грибов. В этом же слое распространены плесневые и слизистые грибы, а также протесты. Все эти организмы, пишет П. Мюллер, оказывают существенное влияние на образование муля, но особенно большая роль принадлежит дождевым червям. Кроме Lumbricus terrestris, встречаются другие виды этого рода, а также представители рода Allolobophora. По свидетельству П. Мюллера, на почве с описанным типом перегноя бук прекрасно растет и возобновляется.
В дальнейшем О, Тамм и другие исследователи показали, что перегной типа «муль» присущ главным образом серым и коричневым лесным почвам, северным (оподзоленным) черноземам, буроземам Раманна. Значительно реже его можно встретить у дерново-подзолистых почв.
В буковых лесах П. Мюллер описал и другой тип перегноя — мор, или грубый перегной. Этот тип перегноя свойствен преимущественно подзолам и многим дерново-подзолистым почвам.
В напочвенном покрове лесов с грубым перегноем получают значительное распространение зеленые блестящие мхи: Hypnum, Polytrichum, Dircanum, к которым обычно присоединяется черника, брусника и некоторые другие. Из травянистых растений развиты преимущественно луговик извилистый (Deschampsia flexuosa), седмичник (Trientalis europaea).
Микроскопический и механический анализ показал, что грубый перегной состоит из буковых листьев, цветков и плодов, переплетенных мельчайшими разветвлениями буковых корней, сплетенных в плотную массу очень прочным грибным мицелием. По определению Рострупа, этот мицелий принадлежит Cladosporium epiphyllum или особому виду, который Роструп предложил назвать Cladosporium humifacilum. Почвы с грубым перегноем отличаются бедностью почвенно-подстилочной фауны, в них лишь изредка попадаются ракообразные и черви из группы Cordiaceae и Anguillulae.
Грубый перегной (мор) имеет вид вязкого, плотного, черно-бурого слоя, который довольно резко граничит с лежащим под ним горизонтом подзола.
Содержание органического вещества от 30 до 50%; оно резко снижается с глубиной. В нем содержится значительное количество водорастворимых органических веществ.
На почвах с грубым перегноем бук растет значительно хуже, естественного возобновления нет, а искусственное сопряжено с большими трудностями.
Между этими двумя крайними типами перегноя имеется серия постепенных переходов. П. Мюллер описывает почвы, обладающие мягким и рыхлым, но лишенным зернистой структуры слоем. Значительное содержание в этом слое хитина насекомых послужило П. Мюллеру основанием назвать этот тип перегноя Insectenmor, исходя из предположения, что он образован в результате разрыхления насекомыми грубого перегноя.
В дальнейшем, как средний тип между грубым перегноем (мор) и мягким (муль), был выделен промежуточный тип модер.
Тип мор был разделен на грубый перегной (Rohhumus) и сухой торф (Trockentorf). Иногда эти два наименования употребляли как равнозначные.
Надлежит особо подчеркнуть, что по наблюдениям Мюллера под одной и той же древесной породой может образоваться как гумус типа муль, так и грубый гумус. Характер образуемого перегноя зависит в большей степени от живого напочвенного покрова и эдафона, чем от состава древесной растительности. П. Мюллер полагал, что изменения в почвенной фауне приводят к изменениям мертвого напочвенного покрова и даже состава древостоя.
В последующих работах немецких исследователей лесных почв развитые П. Мюллером положения претерпели существенные изменения. По мнению И. В. Тюрина «смысл этих понятий был подвергнут искажению».
Е. Раманн, изменив значение термина муль, начал применять его для характеристики части почвы, содержащей экскременты червей, и называть мулем всякую рыхлую лесную почву. В дальнейшем Р. Альберт применял термин муль только для обозначения почв, в которых перегной проникает в нижние слои «в растворенном» виде. Когда органические вещества скапливаются на поверхности почвы в виде неразложившихся растительных остатков, он применял термин модер.
Выделение без должного обоснования многочисленных разновидностей лесного перегноя породили стремление вернуться к простой классификации П. Мюллера.
Л. Роммель и С. Гейберг предложили сохранить разделение перегноя лесных почв на два типа, предложенные П. Мюллером: муль и грубый перегной (мор). Для первого они сохранили старый термин — муль, а для грубого перегноя ввели наименование дэф (duff). В дальнейшем каждый из названных типов они подразделяли па несколько разновидностей: рыхлый муль (Crumb mull), зернистый муль (grain mull), двойной муль (twin mull), измельченный муль (detritus mull), корневой дэф (root duff), лиственный дэф (leaf duff), жирный дэф (greasy duff), волокнистый дэф (fibrous duff).
В последующих исследованиях Р. Фальк (1909) связывал образование двух главных типов лесного перегноя с характером почвенной микрофлоры. Он полагал, что существует два типа разложения растительных остатков в лесных почвах: коррозия и деструкция. Согласно Р. Фальку, при коррозии в равной степени разрушаются клетчатка и лигнин, при деструкции клетчатка разрушается почти полностью, а лигнин, слабо затрагивается разрушением и накапливается. Коррозия приводит к более полной гумификации растительных остатков и образованию перегноя типа муль. При деструкции разложение менее полное, происходит накопление не вполне разложившихся растительных остатков, лишенных клетчатки, но относительно обогащенных лигнином. При этом образуется грубый перегной, или мор (дэф — у американцев).
Процесс коррозии, по исследованиям Р. Фалька, обусловлен деятельностью гриба Agaricus nebularis, обитающего в подстилке березового леса, а также других грибов, свойственных лиственным лесам. Эти грибы обладают способностью разрушать не только клетчатку, но и лигнин. Типичный гриб, вызывающий деструкцию, — Coniophora. Этот гриб разрушает только клетчатку и гемицеллюлозу, а лигнин не затрагивает. Грибы последнего рода преобладают в почвах чистых хвойных лесов. В смешанных хвойно-лиственных лесах в большом количестве встречаются грибы, разрушающие лигнин, что приводит к образованию мягкого перегноя типа муль.
Представляет интерес разработанная Н. Н. Степановым (1929, 1932) классификация лесных подстилок еловых лесов, построенная на принципе, который можно назвать генетическим. В основу этой классификации положен характер того растительного опада, который служит материнским веществом для образования подстилки.
Свойства лесных подстилок находятся в тесной генетической зависимости от состава тех растительных остатков, из которых образуется лесная подстилка, и от условий, в которых идет их гумификация. Поскольку живой напочвенный покров служит довольно хорошим показателем водно-воздушного режима и содержания в почвенном профиле легкоподвижных элементов питания, то разделение лесных подстилок еловых лесов по характеру растительности напочвенного покрова дает представление о материале, служащем для образования лесной подстилки, и об условиях, при которых идет ее образование.
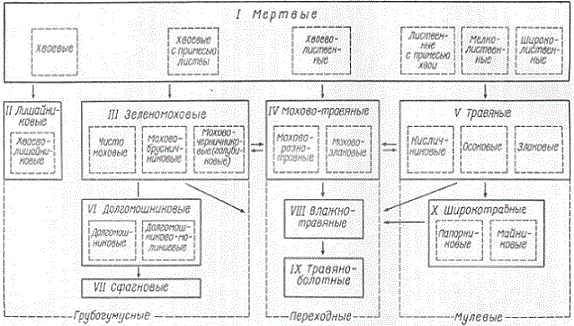
Классификация лесных подстилок (схема Н. Н. Степанова)
Сказанное послужило основанием для распространения принципа, предложенного Н. Н. Степановым для лесных подстилок ельников, на другие типы леса. В результате была предложена общая схема классификации лесных подстилок. На схеме стрелками показаны наиболее распространенные переходы одного вида подстилок в другие. Стрелки обозначают наиболее распространенные переходы между группами подстилок, обведенными сплошными черными линиями, а не между их разновидностями, которые обведены пунктирными линиями.
На основании обобщения исследований немецких и американских ученых Е. Эвальд в 1956 г. предложил следующую схему гумусообразования в лесных почвах:
I. Гумусообразование без влияния грунтовых или застойных вод.
А. Гумусообразование при участии животных:
- земляных червей,
- насекомых.
Б. Гумусообразование при участии грибов.
II. Гумусообразование под влиянием грунтовых и застойных вод.
В основе деления гумусообразования без влияния грунтовых и застойных вод лежат представления Л. Роммеля о влиянии соотношения между развитием микрофлоры и почвенно-подстилочных животных на тип гумусообразования. Л. Роммель пришел к выводу, что если деятельность почвенных животных подавляет развитие грибной флоры, там получает развитие бактериальное разложение мертвых растительных остатков и синтез мягкого перегноя типа муль. В противоположном случае получает преобладание грибное разложение, приводящее к образованию грубого перегноя (мор).
На этом основании Е. Эвальд выделяет два типа: гумусообразование при участии животных и гумусообразование при участии грибов. Дождевые черви измельчают растительные остатки, пропускают их через свой пищеварительный канал, где они подвергаются сложным, превращениям с образованием перегнойных веществ, выделяемых в составе копролитов. Этот тип гумификации присущ почвам с благоприятными влажностью, реакцией почвы, содержанием питательных веществ, образующихся в лесах с легкоразлагаемым опадом, где имеется обильная фауна дождевых червей, с микрофлорой, в которой бактерии и актиномицеты сильно преобладают и грибы не определяют процесса разложения. В этих условиях разложение отмерших растительных остатков происходит быстро с образованием мягкого перегноя.
Там, где условия для дождевых червей неблагоприятны, приобретает значение деятельность насекомых, которые разрушают растительные остатки, перемешивают с почвой, обогащают почву органическим веществом своих экскрементов. Главное отличие заключается в менее полном перемешивании органических веществ с минеральной почвой перегнойного горизонта, состоящего в основном из экскрементов насекомых. Такая форма перегноя соответствует мулеподобному модеру или просто — модеру.
При неблагоприятных условиях в почвах, бедных основаниями и элементами питания, на которых и растительные остатки бедны основаниями, получает преобладание грибной тип разложения. Характерен недостаток оснований для нейтрализации образующихся перегнойных кислот, что приводит к установлению кислой реакции. Как уже было сказано, грибное разложение приводит к образованию грубого перегноя (мор).
Особый характер имеет гумусообразование при заметном влиянии грунтовых или застойных поверхностных вод. Подробные исследования были проведены в черноольшаниках.
Неблагоприятный кислородный режим снижает скорость минерализации мертвых растительных остатков. Низкое содержание кислорода в почве черноольшаников, по-видимому, достаточно для минерализации растительных остатков, но синтезируемые при этом перегнойные вещества оказываются устойчивыми и накапливаются в почве в значительных количествах.
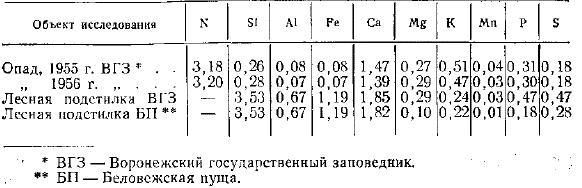
Данные анализа опада и лесной подстилки черноольшаников, (в % на вещество, высушенное при 80—90°) (аналитик Е. Н. Пластинина)
Высокая влажность почв черноольшаников дает основание предполагать, что недостаточная аэрация будет подавлять деятельность нитрофицирующих микроорганизмов при разложении растительных остатков. Исследования показали, что несмотря на высокое содержание аммония, накопление азота нитратов сравнительно невелико. Это показывает, что имеются определенные условия, задерживающие нитрификацию.
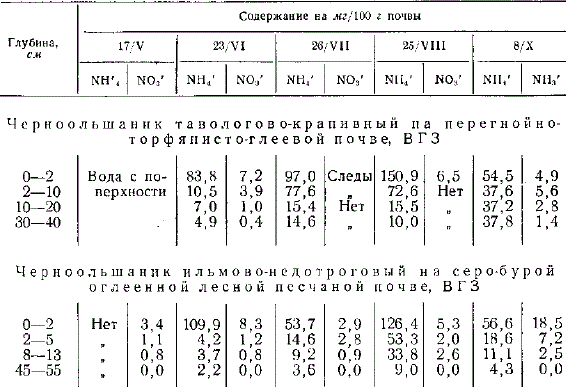
Содержание аммония и нитратов в почвах черноольшаников (аналитик Е. Н. Пластинина)
Недостаток кислорода не может служить единственной причиной подавления нитрификационной активности. Возможно, что отрицательное влияние на процесс нитрификации оказывает сравнительно высокое содержание водорастворимого органического вещества.
В результате своеобразного характера разложения растительных остатков растительность черноольшаников развивается в условиях обильного аммонийно-нитратного азотного питания при резком преобладании аммонийных соединений азота.
При исследовании лесных почв слой лесной подстилки долгое время обозначали знаком А0, а перегнойно-аккумулятивный горизонт знаком А1. Более детальное ознакомление со строением лесных подстилок и подстилко-мохового покрова показало недостаточность такого разделения. Было предложено несколько схем разделения лесной подстилки на слои.
Почва с мягким перегноем (муль) покрыта слоем древесного опада небольшой мощности, который может быть выделен как подстилка. К. Борнебуш называет этот слой «поверхностный муль». Поскольку названный слой состоит преимущественно из опавших листьев, находящихся па различных стадиях разложения, то его обозначают латинской буквой L, начальной буквой английского слова Litter — листва. Непосредственно под ним лежит перегнойно-аккумулятивный горизонт почвы или «истинный муль» (у К. Борнебуша).
В типах леса с хорошо развитым травяным покровом верхний слой почвы густо переплетен корневищами и корнями травянистой растительности, чем напоминает дернину луговых почв.
В почвах с грубым перегноем (мор) можно выделить несколько слоев лесной подстилки. Р. Альберт (1919), Г. Краусс и В. Гроскопф (1928), К. Майвальд (1931) разделяют подстилку на следующие три слоя: 1) свежеопавшие и малоразложившиеся листья, хвоя, ветки, сучья, шишки; 2) грубый перегной (модер, поверхностный торф), представляющий однородную массу, в которой еще можно различить остатки растений, частично сохранивших свой внешний вид, и 3) собственно перегной почвы, в котором перегной тесно связан с минеральной частью и не поддается механическому отделению от минеральной части.
Близкое к изложенному разделение лесной подстилки было предложено Г. Гессельманом (1914). Он выделил также три слоя: 1) свежий лесной опад; 2) слой разложения или ферментации; 3) слой, в котором бесструктурное аморфное органическое вещество тесно соединено с минеральной частью почвы. Первый слой он обозначает буквой L (от слова Laub, Litter), второй слой буквой F по преобладающему процессу ферментации (Fermentation) и третий буквой H от слова Humus.
Американские исследователи также выделяют в подстилке три слоя, но дают им другие наименования: 1) литтер; 2) дэф (duff) и 3) лифмольд (Leafmold).
Н. Н. Степанов, опубликовавший ряд исследований лесных подстилок, также разделял их на три слоя, для обозначения которых в старый знак А0 добавлял порядковые цифры А01, A02, A03. Этого же обозначения придерживается А. П. Малянов.
Рассмотренные способы деления на слои лесных подстилок не имеют принципиальных отличий. Наиболее целесообразными следует признать обозначения, основанные на генетическом принципе Г. Гессельмана. В качестве индексов им взяты начальные буквы наиболее характерных свойств соответствующих слоев. Выделенные три слоя представляют последовательные стадии разложения лесного опада. В 1955 г. на рабочем совещании по картированию лесных типов местообитания было принято подразделение, предложенное Г. Гессельманом.
Значительные работы по изучению морфологии и физических свойств лесных подстилок были проведены лабораторией лесного почвоведения Всесоюзного научно-исследовательского института лесного хозяйства. За ряд лет сотрудниками ВНИИЛХ был собран многочисленный материал по измерению мощности и массы лесной подстилки в различных типах леса европейской части СССР. На основании нескольких тысяч измерений можно сказать, что мощность лесной подстилки в незаболоченных типах леса колеблется в пределах 1—10 см, с преобладанием мощности около 3 см. Масса подстилки изменяется в пределах 14—80 т/га, в заболоченных Типах леса она достигает 120 г/га. В хвойно-моховых незаболоченных лесах преобладает запас около 30 т/га.

Средние данные о колебании мощности и массы лесных подстилок при различных типах почвенного покрова (по материалам лаборатории почвоведения ВНИИЛХ)
Сотрудником той же лаборатории А. П. Малиновым (1937) был проведен вещественный анализ различных слоев лесной подстилки с разделением на хвою, листья, ветки, кору, плоды, гумифицированную часть. На основании полученных материалов А. П. Малиновым сделаны следующие выводы.
В разных типах леса подстилка содержит неодинаковое количество составных частей опада. Состав подстилки меняется в зависимости от строения древостоя, развития подроста и подлеска, возраста, полноты, санитарного состояния и т. д. При переходе к нижним слоям подстилки содержание составных частей опада уменьшается, а содержание продуктов гумификации возрастает. Разложение составных частей подстилки идет с неодинаковой скоростью. Наиболее медленно разлагаются кора и плоды (шишки), что подтверждает относительное увеличение их содержания в нижнем слое подстилки А03. При переходе к нижним слоям подстилки возрастает содержание минеральных частиц, наибольшее количество которых приходится на нижнюю часть, находящуюся на контакте с перегнойно-аккумулятивным горизонтом А1. В этой части содержание минеральных частиц достигает 40%.
Необходимо указать, что мощность и масса лесной подстилки, а также ее вещественный состав отличаются изменчивостью во времени. Поздней осенью, после опада листвы и значительной части хвои, мощность и масса лесной подстилки повышаются, выше становится содержание в ней неизмененных растительных остатков. К концу лета и началу листопада мощность и масса лесной подстилки наименьшая. В весенний и летний период шло интенсивное разложение ее составных частей.
Следует также учитывать большую пространственную неоднородность лесной подстилки, что определяет необходимость при ее исследовании прибегать к большему числу повторностей (А. С. Скородумов, 1939). Эта пространственная неоднородность хорошо видна на прилагаемых картограммах. Особенно велика мозаичность лесных подстилок в хвойных и хвойно-лиственных лесах. В дубравах лесостепи подстилко-травяной покров отличается большой однородностью.
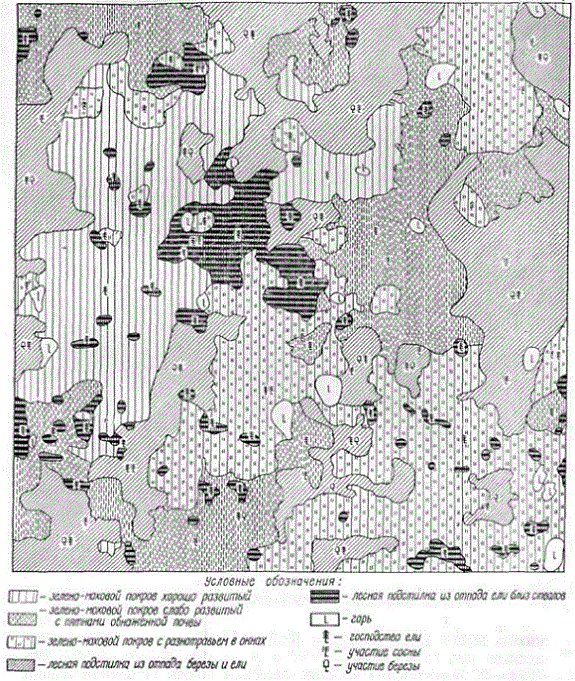
Мозаичность лесной подстилки елового леса

Мозаичность лесной подстилки соснового леса
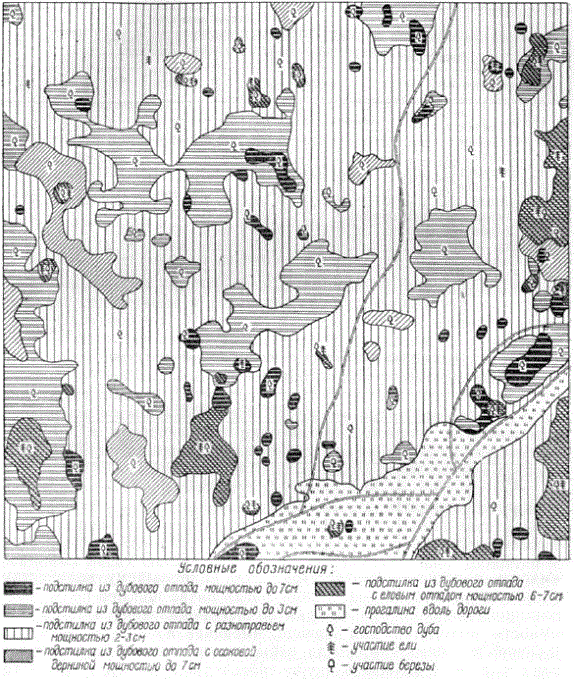
Мозаичность лесной подстилки дубового леса
Исследование физических свойств лесных подстилок показало зависимость степени уплотнения подстилки от режима влажности мест ее образования. Так, подстилка незаболоченных сосняков (1 и 2) из Савальского лесхоза Воронежской обл. имеет объемный вес 0,04—0,05, а в условиях более влажного климата Московской области подстилка сходных сосняков имеет объемный вес 0,07—0,13. В соответствии с этим лесные подстилки имеют очень высокую общую порозность (92,6—97,7), особенно в сухих местообитаниях. При увлажнении нижний слой лесных подстилок сильно набухает. Из всего объема, занимаемого лесной подстилкой, только от 2,3 до 7,4%. приходится на твердую фазу, остальное занято водой и воздухом. Нижние ее слои более уплотнены и порозность в них меньше, чем в верхних слоях. Свободные промежутки в подстилке (ее порозность) заполнены водой и воздухом.
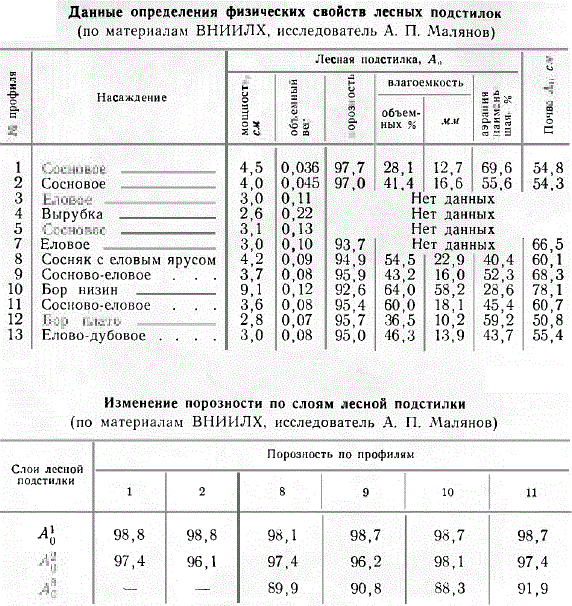
Данные определения физических свойств лесных подстилок и изменение порозности по слоям лесной подстилки
Следует различать: 1) полную влагоемкость подстилки, соответствующую наибольшему количеству воды, удерживаемой подстилкой, когда почва под ней полностью насыщена водой и практически водонепроницаема, и 2) влагоемкость подстилки над почвой, имеющей влажность не выше полевой влагоемкости, когда избыточная влага может из подстилки просачиваться в почву. Последний вид влагоемкости наиболее часто встречается в природных условиях. Полное насыщение подстилки водой наблюдается в незаболоченных почвах лишь в период высокой весенней, а иногда и осенней влажности почвы, а в заболоченных почвах более продолжительное время при близком стоянии почвенно-грунтовых вод. По полученным А. П. Малиновым материалам видно, что полная влагоемкость различных подстилок колеблется от 28 до 64 объемных процентов, или от 10 до 58 мм, возрастая с увеличением плотности подстилки. Одна весовая часть подстилки удерживает 5—7 частей воды.
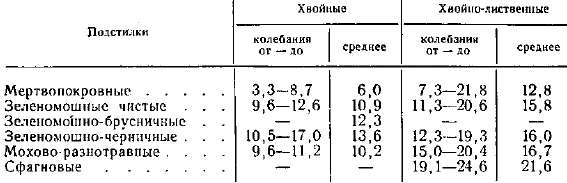
Средние данные водоудерживающей способности подстилко-мохового покрова хвойных и хвойно-лиственных лесов
Насыщающая подстилку вода, заполняя поры, тем самым снижает содержание в подстилке свободного воздуха и затрудняет воздухообмен. Разность между общей величиной порозности и объемом содержащейся в почве воды условно названа аэрацией почвы. Исследования А. П. Малянова показали, что даже в период полной влагоемкости подстилки еще обладают заметной аэрацией (29—70%).
Полное насыщение лесной подстилки водой бывает не часто. Поэтому в последующих работах лаборатории почвоведения ВНИИЛХ внимание было сосредоточено на изучении количества влаги, удерживаемой лесной подстилкой в подвешенном состоянии, т. е. над относительно сухой почвой.
В лиственных лесах были встречены существенные затруднения с определением влагоемкости. На протяжении вегетационного периода запас подстилки в широколиственных, березовых и осиновых лесах сильно колеблется, а следовательно, меняется и влагоемкость. Почвенно-подстилочные животные обогащают лесную подстилку частицами почвы и экскрементами, которые при намачивании набухают, расплываются и вымываются, что отражается на результатах определения.
Определение водоудерживающей способности подстилок в дубравах Шипова леса в 1937 г. дало величины от 9 до 12 мм (среднее из 77 определений), а в 1938 г., поздним летом, 4,8—6,5 мм (среднее из 120 определений).
Более устойчивые и отчетливые результаты были получены при работе в хвойных и хвойно-лиственных лесах. Как правило, водоудерживающая способность в группе чистых хвойных лесов заметно ниже, чем в хвойно-лиственных. В пределах каждой группы подстилок наблюдается повышение водоудерживающей способности при переходе от мертвопокровных к зелено-моховым, а затем сфагновым подстилкам. Следует отметить значительные колебания в пределах каждой разновидности подстилок.
Поскольку подстилко-моховой покров только при продолжительной засухе бывает полностью иссушен, то практически водоудерживающая способность лесных подстилок оказывается соответственно меньше приведенных величин.