К комплементарным, или дополнительным, генам относят такие гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состояниях (А—В—) обусловливают развитие нового признака.
Действие же каждого гена в отдельности (А—bb или ааВ—) воспроизводит признак лишь одного из скрещиваемых родителей.
Впервые такого рода взаимодействие было обнаружено у душистого горошка (Lathyrus odoratus). При скрещивании двух рас этого растения с белыми цветками у гибрида F1 цветки оказались пурпурными. При самоопылении растений F1 в F2 наблюдалось расщепление по окраске цветков в отношении, близком к 9:7. Один фенотипический класс (9/16) имел такую же окраску цветков, как и у растений первого поколения, а второй (7/16) — белую окраску, такую же, как у родительских растений.
Чтобы выяснить, укладывается ли это расщепление в схему дигибридного менделевского расщепления, представим себе, что у каждой исходной расы душистого горошка имеется в гомозиготном состоянии лишь по одной из доминантных аллелей (AAbb и ааВВ), которые при взаимодействии определяют развитие окраски. Поскольку у гибрида первого поколения присутствуют доминантные аллели обоих генов (АаВb), цветки гибридных растений F1 будут окрашенными. Во втором поколении происходит расщепление в отношении 9/16А—В—: 3/16А—bb : 3/16ааВ—: 1/16aabb. Каждый из генов в отдельности не может обусловить развитие окраски, так как выработка антоциановых пигментов осуществляется лишь при наличии доминантных аллелей обоих генов. Поэтому растения с генотипами А—bb, ааВ— и aabb имеют белые цветки и во втором поколении наблюдается расщепление по фенотипу в отношении 9 : 7. Анализирующим скрещиванием и анализом в F3 можно точно подтвердить данное выше объяснение.
Приведем еще несколько примеров, иллюстрирующих действие комплементарных генов у растений и животных.
У земляники развитие «усов», т. е. вегетативных самоукореняющихся побегов, определяется доминантной аллелью, а «безусость» — рецессивной. Но существуют такие формы безусой земляники, которые при скрещивании друг с другом дают гибрид F1 с сильно выраженным признаком «усатости». Исследованиями Т. С. Фадеевой было показано, что в потомстве такого гибрида в F2 получается расщепление, близкое к отношению 9 : 7, а именно: из 752 растений F2 419 оказались с усами, 333 — без усов. Это соответствует теоретически ожидаемому расщеплению: 752 X 9/16 = 423 и 752 X 7/16 = 329.
У белого клевера имеются формы с высоким и низким содержанием цианида. Цианиды, как известно, блокируют дыхательный фермент, но повышают активность папаина (растительной протеазы), катепсина и других ферментов. Высокое содержание цианида в белом клевере связано с усиленным вегетативным ростом без снижения его кормовых качеств. При скрещивании растений с высоким и низким содержанием цианида в F1 доминирует первое свойство, а в F2 наблюдается расщепление, близкое к отношению 3 : 1.
Эти результаты указывают на то, что в данном случае альтернативные признаки определяются одной парой аллелей. Но иногда при скрещивании двух растений клевера с низким содержанием цианида гибриды F1 характеризуются высоким его содержанием, а в F2 расщепление оказывается близким к отношению: 9/16 — с высоким содержанием цианида и 7/16 — с низким. Так же, как у душистого горошка, в данном случае имеет место обычное дигибридное расщепление, в котором 9/16 потомков обладают двумя доминантными генами А—В—, а 7/16 относятся к трем остальным фенотипически неотличимым классам: 3/l6A—bb + 3/16ааВ + 1/16aabb = 7/16. Доминантные аллели разных генов в отдельности не увеличивают содержание цианида по сравнению с тем низким уровнем, который характерен для растения, гомозиготного по рецессивным аллелям обоих генов, но при совместном действии доминантных аллелей обоих генов содержание цианида повышается.
Подобное явление можно показать на примере кукурузы. При скрещивании некоторых форм кукурузы с белыми зернами в F1 зерна в початках оказываются пурпурными. В F2 происходит расщепление на 9/16 пурпурных (А—В—) и 7/16 белых (ааВ—, А—bb и aabb).
До сих пор мы рассматривали примеры комплементарного взаимодействия доминантных генов, при котором каждый из генов в отдельности не обладал способностью вызывать развитие признака. Последний развивался лишь в результате взаимодействия доминантных аллелей двух генов. В силу этого в F2 обнаруживались только два фенотипических класса в соотношении 9:7. Известны, однако, случаи, когда один или оба комплементарных гена характеризуются самостоятельным проявлением. В соответствии с этим меняется и характер расщепления в F2.
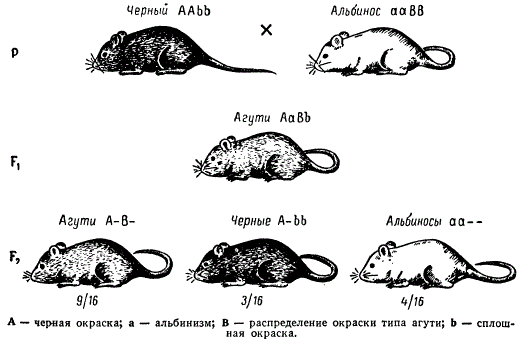
Рассмотрим наследование трех типов окраски шерсти у мышей: дикой, или рыжевато-серой (агути), черной и белой. Окраска дикого типа зависит от наличия гена, определяющего развитие окраски, и от гена, обусловливающего распределение пигмента по длине волоса. Каждый волос у мышей агути имеет по длине кольцо желтого пигмента, а в основании и на конце волоска — черный пигмент. Такое зонарное распределение пигментов и создает окраску агути, свойственную диким грызунам (белка, кролик, морская свинка и др.). У черных мышей отсутствует зонарное распределение пигмента — волосы до всей длине окрашены равномерно. Белые мыши с красной радужной оболочкой глаз, так называемые альбиносы, лишены пигмента.
Надо сказать, что альбинизм встречается у животных почти всех классов — млекопитающих, птиц, амфибий и др. Встречается альбинизм и у человека. Так, например, иногда у родителей-негров рождаются дети альбиносы, т. е. с белой кожей и белыми волосами, но с чертами лица негритянского типа. Известны случаи, когда в семье негров рождаются двойни (разнояйцевые), и один из детей оказывается альбиносом. Такой ребенок имеет рецессивную аллель гена альбинизма в гомозиготном состоянии.
Окраска шерсти у мышей типа агути доминирует над черной, и над белой. При скрещивании черных мышей с белыми с белыми, все гибриды F1 оказываются агути, а в F2 наблюдается расщепление в отношении 9/16 агути : 3/16 черных : 4/16 белых.
Наследование окраски у мышей при взаимодействии двух пар генов (комплементарность)
Взятые в скрещивание мыши-альбиносы являются, очевидно, гомозиготными по рецессивной аллели гена окраски и доминантной аллели гена попарного распределения пигмента (ааВB), а черные мыши — гомозиготными по доминантной аллели гена окраски и рецессивной аллели гена распределения пигмента в волоске (AAbb). У гибридов F1 (АаВb) вследствие взаимодействия доминантных аллелей обоих генов развивается окраска типа агути. Такая же окраска характерна и для 9/16 особей в F2 с генотипом А—В—. Черными в F. оказываются мыши, имеющие генотип А—bb, а белыми — все остальные — (ааВ— и aabb) в силу отсутствия у них гена А, определяющего образование пигмента. Ген В в отсутствие гена А не имеет собственного проявления.
Подобные примеры наследования известны и у растений (лук, кукуруза и др.). У лука скрещивание формы, имеющей неокрашенную (белую) луковицу, с формой, имеющей желтую луковицу, дает в F1 растения с красными луковицами, а в F2 появляются растении с красными (9/16), желтыми (3/16) и белыми (4/16) луковицами. В этом случае опять-таки одна из доминантных аллелей двух генов способна действовать самостоятельно (определяет желтую окраску луковицы), а другой ген проявляется лишь в присутствии комплементарного гена.
Известны и такие случаи, когда каждый из двух комплементарных генов способен проявлять свое действие самостоятельно. Один таких примеров мы уже рассматривали при анализе наследования формы гребня у кур. Каждая из доминантных аллелей генов обусловливала развитие гребня определенной формы (гороховидной или розовидной), а взаимодействие этих генов определяло развитие новой формы гребня ореховидной. В данном примере каждый из комплементарных доминантных генов характеризуется собственным специфическим эффектом, а взаимодействие между ними приводит к новообразованию, к новому выражению признака.
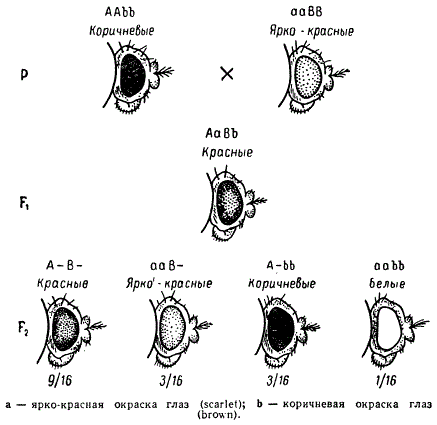
Ряд подобных примеров наследования известен и у других животных и растений. Так, у дрозофилы рецессивная аллель гена scarlet в гомозиготном состоянии определяет ярко-красную окраску глаз, а рецессивная аллель другого гена — brown (также в гомозиготном состоянии) определяет коричневую окраску глаз. При скрещивании гибриды F1 оказываются красноглазыми (дикого типа), рели же оба эти рецессивных гена находятся в гомозиготном состоянии, то такая особь оказывается белоглазой. Если скрестить красноглазых мух F1 друг с другом, то во втором поколении по признаку окраски глаз будет наблюдаться расщепление на 4 фенотипических класса в отношении 9/16 красных : 3/16 ярко-красных : 3/16 коричневых : 1/16 белых. Такое поведение признаков в наследовании также говорит о расщеплении по двум комплементарным генам с самостоятельным действием.
Наследование окраски глаз у Drosophila при взаимодействии двух пар генов (комплементарность)
Если генотип мух с коричневыми глазами условно обозначить ААbb, с ярко-красными — ааВВ, а генотип красноглазых гибридов F1 — АаВb и белоглазых мух — aabb, то фенотипические радикалы полученных в F2 классов могут быть представлены как А—В— (9/16), ааВ— (3/16), А—bb(3/16) и aabb(1/16).
Природа взаимодействия генов в этом случае более ясна, чем в случае наследования формы гребней у кур. Нормальная красная окраска глаз у мух обеспечивается в основном тремя видами пигментов красным, коричневым и желтым. В гомозиготном состоянии рецессивный ген а блокирует образование коричневого пигмента, вследствие чего развиваются ярко-красные глаза, а другой рецессивный ген b в гомозиготном состоянии блокирует одновременно образование красного и желтого пигментов, и поэтому развиваются коричневые глаза. В F1 объединяются доминантные аллели этих генов, поэтому образуются все пигменты, дающие в совокупности красную окраску глаз. Новый класс белоглазых мух, появляющихся в F2, очевидно, является результатом одновременного блокирования синтеза всех трех пигментов.
Подобные примеры можно привести и на растительных объектах. Известно, что окраска плодов у томатов обусловливается каротиновыми пигментами (ликопины и бета-каротин), имеющими огромное значение в синтезе витаминов. Анализ наследования окраски плодов у томатов показывает, что красная окраска плодов определяется взаимодействием комплементарных доминантных генов R и Т, оранжевые плоды образуются на растениях с генотипом R—tt, желтые — на растениях с генотипом rrТ—, промежуточные, желтооранжевые — на растениях rrtt. Здесь также расщепление в F2 соответствует генетической формуле дигибридного скрещивания 9:3:3: 1. При этом установлено, что красные и оранжевые плоды содержат наибольшее количество каротинов, а желтые — наименьшее. Двойной рецессив содержит промежуточное количество каротинов в плоде. Качественные различия в наборе каротинов соответствуют определенным различиям в генотипе.
Во всех разобранных примерах мы имели дело с комплементарным взаимодействием доминантных и рецессивных неаллельных генов. Взаимодействие доминантных генов обусловливало развитие ореховидного гребня у кур, красной окраски глаз у дрозофилы, красной окраски плодов у томатов. Взаимодействие рецессивных аллелей этих генов приводило к развитию пластинчатого, или ликвидного, гребня у кур, белых глаз у дрозофилы, желто-оранжевой окраски плодов у томатов.
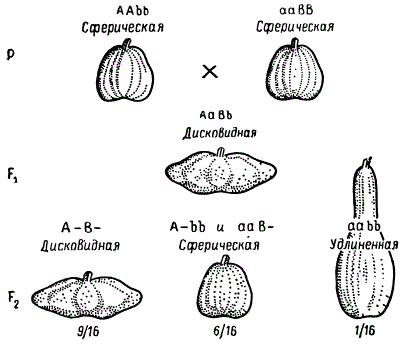
Следует отметить, что в ряде случаев комплементарные гены, способные к самостоятельному проявлению, при отсутствии дополнительного гена могут давать каждый в отдельности сходный фенотипический эффект. Характер расщепления дигибрида в F2 при этом также изменяется. Так, у тыквы (Cucurbita реро) имеются сорта с разной формой плода: сферической, дисковидной и удлиненной. Сферическая форма плода является рецессивной по отношению к дисковидной.
Наследование формы плода у Cucurbita pepo при взаимодействии двух пар генов (комплементарность)
От скрещивания растений с плодами сферической формы, но имеющими разное происхождение, возникают гибридные растения, дающие плоды только дисковидной формы. В потомстве этих растений в F2 появляются три фенотипических класса в отношении 9/16 дисковидными плодами, 6/16 — со сферическими и 1/16 — с удлиненными. Зная закономерности дигибридного расщепления при взаимодействии генов, нетрудно понять, что и здесь имеет место взаимодействие двух генов, влияющих на развитие формы плода, каждый из доминантных комплементарных генов обусловливает развитие плодов сферической формы, а их взаимодействие приводит к образованию дисковидных плодов. Взаимодействие рецессивных аллелей этих генов определяет развитие плодов удлиненной формы.
Рассматривая примеры комплементарного действия генов, мы убеждаемся, что такое взаимодействие генов приводит к развитию признаков, свойственных диким предкам данных видов (серая окраска грызунов, дисковидная форма у тыквы и т. д.). Некоторые авторы рассматривают это явление как пример атавизма. Эти представления основываются на предположении, что в процессе эволюции животных и растений доминантные гены, действующие комплементарным образом, изменились, мутировали в рецессивное состояние (А → а, В → b, С → с и т. д.).
У диких предков домашних животных и растений доминантные гены комплементарного действия поддерживались естественным отбором вместе в одном генотипе (например, серая окраска грызунов, дисковидная форма плода у тыквы, красная окраска глаз у дрозофилы и др.). При одомашнивании и проведении селекции с помощью скрещиваний и искусственного отбора комплементарные гены как бы разобщились. Генотип АаВb разлагался селекционерами на генотипы ААЬЬ и ааВВ. Поэтому при скрещивании таких организмов иногда наблюдается как бы возврат к признакам диких предков.
Мы остановились более подробно на комплементарном действии генов потому, что этот тип взаимодействия иллюстрирует один из путей возникновения комбинативной изменчивости и имеет отношение к широко используемому явлению гибридной мощности — гетерозису.