С помощью трансформации и трансдукции осуществляется односторонний обмен наследственными факторами между бактериями. И эти процессы в какой-то мере компенсируют отсутствие у них настоящего полового процесса.
Поиски полового процесса у бактерий в течение длительного времени были безуспешными. Лишь после того, как были разработаны методы селективных сред и получены штаммы биохимических мутантов, Дж. Ледербергу и Е. Татуму удалось в 1946 г. доказать наличие своеобразного полового процесса у Escherichia coli на примере штамма К12. Процесс переноса генетической информации от одной бактерии к другой при контакте клеток получил название конъюгации.
Рекомбинация у бактерий
Вначале доказательство наличия полового процесса у бактерий основывалось на генетическом методе получения рекомбинантов.
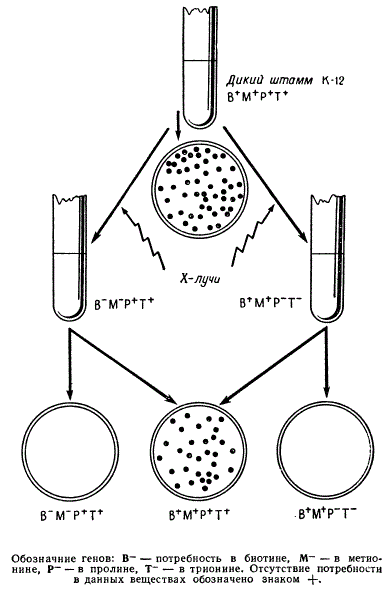
У Е. coli прототрофного штамма K12 был получен ряд биохимических мутантов с различными потребностями в биотине (В—), метионине (М—), пролине (Р—), треонине (Т—) и др. Все штаммы бактерий с этими мутациями не росли на минимальной среде.
Чтобы проверить возможность образования рекомбинантов, были взяты для штамма, различающиеся по генотипу: — В—М—Р+Т+ и В+М+Р—Т— и фенотипу — потребностях в аминокислотах и витамине. Первый для своего роста нуждался в биотине и метионине, но не нуждался в пролине и треонине. Второй штамм нуждался в двух последних веществах и не нуждался в двух первых. Клетки обоих ауксотрофных штаммов в течение некоторого времени выращивали в смешанной культуре, а затем высевали на минимальную среду. Ни один из двух исходных штаммов не мог расти на этой среде. Однако на каждые 109 посеянных клеток из смешанной культуры на минимальной среде вырастало около 100 колоний. Отсутствие потребности в ростовых веществах свидетельствовало о том, что по своему генотипу эти клоны должны были быть В+М+Р+Т+.
Рекомбинация у бактерий
Появление этих колоний нельзя было объяснить трансформацией или трансдукцией, так как рекомбинацию не удавалось обнаружить, если суспензии двух исходных штаммов разделяли бактериальным фильтром. Оставалось предположение, что для появления рекомбинантов необходим непосредственный контакт между бактериальными клетками, при котором происходит обмен наследственным материалом. Поэтому исследователи пришли к заключению, что появление указанных колоний на минимальной среде могло быть следствием осуществления рекомбинации между штаммами бактерий В—М—Р+Т+ и В+М+Р—Т—. Только в результате нового сочетания генов могли появиться клетки с генотипом дикого типа В+М+Р+Т+, растущие на минимальной среде.
Таким образом, на основе чисто генетических методов было сделано предположение о наличии специфического полового процесса у бактерий. Позже с помощью электронной микроскопии были Получены фотографии конъюгирующих бактерий, соединенных попарно тонким протоплазматическим мостиком.
Изучение конъюгации цитологическими методами показало, что анастомозы образуются на ограниченном участке поверхности бактерий, ширина мостика достигает 300 ммк.
Половые типы у бактерий
Изучение ряда штаммов кишечной палочки выявило наличие у нее «половой» дифференциации.
Выражалось это в том, что исследованные штаммы первоначально распались на две группы (типы).
В первой группе штаммов конъюгации клеток не наблюдалось. Во второй группе конъюгация происходила, но количество рекомбинантов было небольшим. В то же время при конъюгации бактерий из разных групп рекомбинанты появлялись в 100—1000 раз чаще.
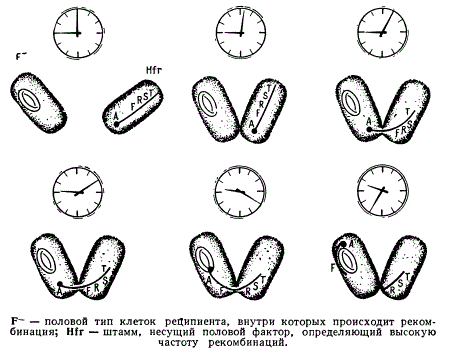
Это позволило рассматривать указанные группы как различные половые типы, которые были обозначены F+ и F—. Скрещивание бактерий штаммов F— x F+ всегда безуспешно, a F+ х F+ лишь редко дает рекомбинантов.
Сравнение штаммов F— и F+ показало, что поведение бактерий этих штаммов в скрещивании функционально различно. Наиболее наглядно это выявилось в опыте, в котором под микроскопом удалось наблюдать пары конъюгирующих бактерий и выделить их с помощью микроманипулятора. В опыт были взяты штаммы F— и F+, клетки которых различались еще и по форме. Затем бактерии разъединяли и изучали потомство каждой из них. Оказалось, что в потомстве клетки F+, участвовавшей в конъюгации, никогда не появляются рекомбинанты. В то же время клетки F— в значительном числе случаев при делениях давали рекомбинанты, совмещающие признаки обоих родителей. Оказалось, что клетка F+ как бы оплодотворяется, являясь «женской», а клетка F+ выступает в роли оплодотворяющей, «мужской». При конъюгации клеток происходит односторонняя передача генетического материала от клетки F+ в клетку F—. При этом клетки F+ являются донорами генетического материала, а клетки F— — реципиентом, и рекомбинации происходят внутри последней.
В скрещиваниях F— x F+ рекомбинанты появляются с частотой 1 на 104 родительских клеток; при этом наблюдается, что очень многие клетки «женского» родителя приобретают характеристику не получая каких-либо других признаков от второй культуры. Передача полового типа происходит независимо от передачи других генетических маркеров.
Позже среди других штаммов кишечной палочки был выявлен и третий половой тип, определяющий высокую частоту рекомбинаций и поэтому обозначенный Hfr (High frequency of recombination). Клетки этого типа возникают мутационно в культурах F+. Скрещивания F— х Hfr дают особенно высокий процент рекомбинантов: 1 на 10 исходных клеток. Вместе с тем, в отличие от скрещиваний «женские» клетки в скрещивании F— x Hfr не приобретают Свойства F+.
Скрещивания F— х F+ показывают, что фактор фертильности (F-фактор), обусловливающий принадлежность к «мужскому» типу передается с высокой частотой независимо от остальных генов (автономно). F-фактор в клетках F+ оказывается внехромосомным и в этом отношении ведет себя подобно цитоплазматической частице. Клетки Hfr теряют способность к автономной передаче F-фактора. Среди рекомбинантов в потомстве от скрещивания изредка важно обнаружить клетки Hfr. Генетический анализ показал, что в данном явлении фактор фертильности передается сцепленно с другими генами и занимает определенный локус в бактериальной хромосоме. Таким образом, F-фактор, если он присутствует в клетке, т себя двояко: как автономная цитоплазматическая частица в клетках F+) или как локус хромосомы (в клетках Hfr). Поскольку клетки Hfr возникают из клеток F+, то следует предполагать, что F-фактор может переходить из цитоплазмы на бактериальную хромосому; при этом исчезает способность к его автономной передаче в F—-клетки. Сама «мужская» клетка изменяется так, что в скрещиваниях, передавая своему партнеру хромосомный материал, начинает производить рекомбинанты примерно в 1000 раз чаще, чем клетки F+. В. Хейс установил, что штаммы Hfr являются мутантами Р+-бактерий. Мутации происходят в направлении F+ — Hfr, и это изменение не связано с утратой фактора F, так как при обратном мутировании Hfr → F+ свойство донорства полового фактора восстанавливается.
У бактерий было обнаружено еще несколько генетических детерминантов, ведущих себя подобно F-фактору и способных находиться в клетке в двух альтернативных состояниях — в виде цитоплазматической частицы или в виде локуса бактериальной хромосомы. Для их обозначения потребовалось введение специального термина. Такие детерминанты Ф. Жакоб и И. Вольман назвали эписомами. Под эписомами понимают генетические факторы, которые могут присутствовать в клетке или в виде автономных цитоплазматических частиц, или могут включаться, наподобие части генома профага, в бактериальную хромосому, обнаруживая тогда сцепление с остальными генами или могут совсем отсутствовать в клетке. К эписомам могут быть отнесены и умеренные бактериофаги.
Автономность передачи полового фактора F независимо от генома бактерий является чертой, характерной для эписом. Этот тип передачи напоминает передачу генов при трансдукции, хотя различия между передачей F-фактора и трансдукцией существенны и заключаются в том, что в первом случае для передачи F-фактора необходим контакт между F— и F+-клетками, а во втором — контакт не обязателен.
Генетические карты бактерий
Для картирования генов у кишечной палочки Ф. Жакоб и И. Вольман разработали особый метод.
Из смешанной культуры двух конъюгирующих линий, маркированных теми или иными генами, через разные промежутки времени после начала конъюгации брали порции этой культуры и помещали в гомогенизатор, в котором с помощью механического встряхивания удается разъединить конъюгирующие бактерии. После этого клетки из культуры рассевали на селективные среды, позволяющие выявлять колонии рекомбинантов.
Описанным приемом удалось установить очень интересное явление. Оказалось, что количество наследственного материала, перемещающегося из одной клетки в другую, пропорционально времени конъюгации клеток. Передача всех учитывающихся в группе сцепления маркеров начиналась через 8 мин и заканчивалась позднее, чем через час после начала конъюгации. Поскольку для перемещения разных генов из одной клетки в другую необходимо разное время, то время передачи фрагмента хромосомы «мужских» клеток в «женские» в этом случае может служить мерой расстояния между генами.
Зависимость передачи генетического материала от продолжительности конъюгации у Escherichia coli
В этих опытах был обнаружен и другой очень важный факт, а именно, что единственная группа сцепления Escherichia coli представлена в виде замкнутого круга; она состоит из двунитчатой ДНК длиной 1,2—1,4 ммк.
Различные линии Hfr начинают передачу генов с разных участков хромосомы и в разной последовательности, но линейный порядок генов остается при этом постоянным. Наблюдаемые изменения в последовательности передачи факторов навели на мысль, что у Е. coli в клетках F+ имеется лишь одна, причем кольцевая, группа сцепления. При возникновении клеток Hfr фактор F в разных линиях садится в различных точках кольцевой хромосомы, раскрытие кольца может произойти справа или слева от него. Место разрыва кольца и определяет направление передачи генов характерной для данной линии последовательности. Передача начинается с раскрытого конца кольцевой хромосомы, а на противоположном конце хромосомы всегда оказывается фактор F. Таким образом, свободный от фактора F конец хромосомы оказывается начальной точкой переноса группы сцепления, обозначаемой как локус О (от слова origin). Гены, вошедшие при конъюгации в F-клетку, включаются в ее хромосому посредством процесса, по-видимому, аналогичного кроссинговеру, так как при делении такой «оплодотворенной» клетки появляются рекомбинанты.

Порядок передачи наследственных факторов различными линиями Hfr Escherichia coli
Большинство штаммов Hfr передает не все гены группы сцепления, а лишь какой-то фрагмент бактериальной хромосомы. В последнее время были описаны штаммы, способные к передаче всей группы сцепления. Вся бактериальная хромосома переходит при этом в клетку за 111 мин.
Предполагается, что репликация ДНК начинается с определенной точки кольцевой хромосомы, распространяется по ней и заканчивается у начала. Пока не закончится полностью репликация всей хромосомы, новый цикл не начинается. Репликация согласуется с ростом клетки.
Ф. Жакоб предполагает, что генетический аппарат бактерии состоит из нескольких независимых структур — групп генов, включенных либо в хромосому бактерии, либо в половой фактор, либо в профаг умеренного фага, содержащих по одной молекуле ДНК разной длины. Каждая из таких целых структурных единиц обладает автономной способностью к репликации. Структурную единицу, способную к автономной репликации, определяемой ростом клетки, предложено называть репликоном.
Допускается, что репликоны (например, часть хромосомы и эписомный фактор) прикрепляются к клеточной мембране. Именно этим объясняется, что их репликация связана с ростом мембраны, которая в свою очередь связана с ростом и делением клетки.
Помимо гибридизации между различными штаммами Е. coli, сейчас осуществлена гибридизация между Е. coli и несколькими штаммами тифозных и дизентирийных бактерий Shigella и Salmonella. Данная область исследований обещает быть интересной, так как при этом вскрываются филогенетические отношения различных групп бактерий и выясняются пути происхождения новых патогенных форм кишечных бактерий.
После открытия полового процесса у Е. coli были начаты исследования по гибридизации других бактерий. Получены гибриды между штаммами внутри видов Pseudomonas aeruginosa, Vibrio cholerae, внутри нескольких видов рода Salmonella и др.
Таким образом, у микроорганизмов известны две уникальные особенности «полового» процесса: односторонность передачи генетического материала и ориентированный характер этой передачи.
Возможность составления генетической карты для того или иного организма представляет собой высшее достижение генетического анализа для данного объекта. Генетические карты необходимы для изучения генетического, потенциала вида и тонкого анализа наследственности. К настоящему времени на основе использования прерываемой конъюгации, трансдукции и других способов рекомбинации у бактерий, особенно кишечной группы, хорошо разработаны генетические карты. Недавно М. Демерец подвел некоторые итоги по сравнительному изучению генетических карт у кишечных бактерий и представил детальную карту для Salmonella typhimurium.
Генетическая карта сальмонеллы включает 133 локуса. При этом место того или иного локуса в кольцевой хромосоме определяется интервалом времени, при котором данный локус передается от клетки донора реципиенту при прерываемой конъюгации. Все 133 локуса хромосомы сальмонеллы передаются за 138 мин конъюгации. В случае для кишечной палочки установлено 100 локусов, которые передаются при конъюгации более чем за 100 мин.
На карте видны участки, в которых несколько генов оказываются тесно сцепленными и объединяются общей активностью в биосинтезе веществ. Например, участок биосинтеза пиримидина составляет 9 генов, гистидина — 10 генов, и т. д. Все эти блоки генов чаще всего относятся к одному оперону.
Сравнительное изучение генетических карт показывает, что из 133 генов Salmonella typhimurium 59 генов являются общими с Escherichia coli. При этом они имеют одинаковые функции, т. е. Контролируют сходные этапы биосинтеза и расположены на генетических картах в одинаковой последовательности. Таким образом, между двумя родами бактерий обнаруживается высокая гомология генетических структур. Однако внутригенные различия на молекулярном уровне между этими родами довольно высокие.
Дальнейший сравнительный анализ генетических карт облегчит понимание не только структуры генотипа, но и раскроет путь к молекулярному анализу эволюции генетической системы.