Методы филогенетических исследований органически связаны с методами изучения фактов эволюции. Основным методом филогенетических исследований до настоящего времени нужно считать морфологический метод, так как превращения формы организма остаются наиболее очевидным фактом и позволяют с большой долей успеха проследить явления трансформации видов.
Из этого не следует, конечно, что к филогенетическим исследованиям не приложимы другие методы — физиологический, экологический, генетический и т. п. Форма и функция организма неразрывно связаны. Любой организм слагается под влиянием конкретных факторов среды, он взаимодействует с ней, он находится в определенных связях с другими организмами. Однако форма организма, его строение остается всегда чувствительным показателем всех этих связей и служит руководящей нитью для исследователя вопросов филогенетики. Морфологический метод исследования занимает ведущее положение в изучении филогении и выводы его в общем нашли подтверждение при проверке их другими методами. Большим достоинством морфологического метода является доступность сочетания его со сравнительным методом исследования, без которого нельзя обнаружить самый факт трансформации живых систем. Действительность морфологического метода в высокой степени усиливается тем обстоятельством, что он по существу глубоко самокритичен, так как может быть применен в различных направлениях.
Если мы располагаем большим палеонтологическим материалом (например, эволюция лошади), мы можем применить сравнительно-морфологический метод к последовательным рядам предков и потомков и таким образом выявить направления и способы эволюции данной группы. Рисунок дает представление о существе сравнительно-морфологического метода в применении к предкам лошади. Последовательная редукция боковых пальцев и развитие среднего (III) пальца показывают направление эволюционного развития «лошадинного ряда».
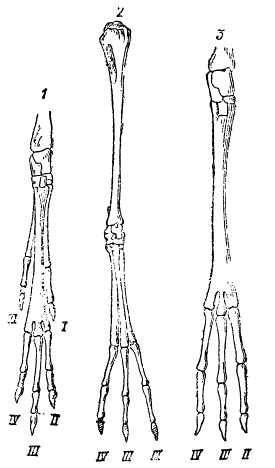
Сопоставление конечностей тушканчиков с убывающим числом пальцев и нарастающей специализацией. 1 — малый тушканчик Allactaga elator, 2 — Salpingotas Koslovi, 3 — мохноногий Dipus sagitta. I—V — пальцы от первого до пятого (По Виноградову)
Далее, палеонтологические данные гармонируют со сравнительно-анатомическими исследованиями современных форм. На рисунке сопоставлены конечности трех форм с убывающим числом пальцев. Хотя это не филогенетический ряд, все же создается представление, что все три конечности являются результатом проявления сходных процессов, достигших разных ступеней развития. Поэтому сравнительно-морфологический метод и по отношению к современным формам, независимо от палеонтологии, делает вероятным предположение, что, например, однопалая нога должна была развиться из многопалой. Когда к этим выводам присоединяются факты сравнительной эмбриологии, показывающие, что, например, у эмбриона лошади боковые пальцы закладываются, а затем они постепенно редуцируются, то наше заключение о происхождении однопалой лошади от многопалого предка становится еще вероятнее.
Совпадение этих данных показывает, что факты палеонтологии, сравнительной анатомии взрослых форм и сравнительной эмбриологии взаимно контролируют и дополняют друг друга, образуя в своей совокупности синтетический тройной метод филогенетических исследований, предложенный Геккелем (1899) и не потерявший своего значения и ныне. Принимается, что совпадение данных палеонтологии, сравнительной анатомии и эмбриологии, в известной мере, служит доказательством правильности филогенетических построений.
Таковы самые общие принципы филогенетических исследований.
Рассмотрим теперь коротко описанные элементы единого метода филогенетических исследований.
Данные палеонтологии наиболее убедительны. Однако они обладают крупным дефектом, а именно, палеонтолог имеет дело только с морфологическими признаками и притом неполными. Организм, как целое, находится за пределами палеонтологического исследования. Ввиду этого палеонтологу особенно важно учесть все доступные для него признаки животных, с останками которых он имеет дело. В противном случае его филогенетические выводы могут оказаться ошибочными.
Положим, что формы А, Б, В, Г, Д, Е сменяют друг друга в последовательных геологических горизонтах, и что палеонтолог имеет возможность наблюдать определенную сумму их признаков — а, б, в и т. д. Допустим, далее, что форма А обладает признаками а1, б1, в1, а у форм Б, В, Г,… эти признаки изменены (соответственно а2, б2, в2.. а3, б3, в3…, ит. д.). Тогда во времени мы получаем такой ряд данных
| Эпохи | Виды | Их признаки |
| Современность | Е | а6 б6 в6 |
| Плейстоцен | Д | а5 б5 в5 |
| Плиоцен | Г | а4 б4 в4 |
| Миоцен | В | а3 б3 в3 |
| Олигоцен | Б | а2 б2 в2 |
| Эоцен | А | а1 б1 в1 |
Этой табличке соответствует, например, «ряд» лошадиных предков, где от эопиппуса до лошади мы имеем преемственную смену в развитии ряда признаков. В таблице показано преемственное развитие по всем ведущим признакам. Каждый последующий признак (например, а4) выводится из каждого предыдущего (например, а3). В подобных случаях становится вероятным, что ряд А, Б, В, Г, Д, Е образует филогенетический ряд, т. е. ряд предков и их потомков. Таков ряд от эогиппуса до лошади и некоторые другие.
Предположим теперь, что мы имеем дело со следующими данными,
| Эпохи | Виды | Их признаки |
| Современность | Е | а1 б5 в1 |
| Плейстоцен | Д | а3 б4 в2 |
| Плиоцен | Г | а2 б3 в1 |
| Миоцен | В | а2 б3 в3 |
| Олигоцен | Б | а4 б2 в2 |
| Эоцен | А | а1 б1 в1 |
т. е. мы констатируем ряд форм, которые последовательно сменяют друг друга во времени, причем по одному из признаков (б) мы получаем картину последовательного развития от б1 до б5. Все же наш ряд не есть филогенетический ряд, так как, например, в отношении признаков а и b мы не наблюдаем последовательной специализации. Например, вид А имеет формулу А (а1, б1, б1), но вид Б явно не является его прямым потомком, так как имеет формулу Б (а4, б2, б2) и т. д. Очевидно, мы здесь имеем дело с последовательными «обломками» филогенетического древа, многие ветви которого не найдены. Поэтому ряд А, Б, В, Г, Д, Е в действительности равен А, Б1, В2, Г3, Д1. Такой ряд называется ступенчатым. Для разъяснения различий между ним и филогенетическим рядом воспользуемся рисунком, показывающим эволюцию лошади. Здесь филогенетическим будет следующий ряд: эогиппус, орогиппус, мезогиппус, парагиппус, меригиппус, плиогиппус, плезиппус, лошадь. Ступенчатым был бы, например, следующий ряд форм: гиракотерий, эпигиппус, миогиппус, анхитерий, гиппарион, гиппидиум, лошадь. Все это не предки и потомки, а последовательные, но разрозненные боковые ветви филогенетического древа.
Как видно, ступенчатый ряд имеет большое рабочее значение, так как, исходя и из него, можно было бы сделать вывод, что лошадь произошла от многопалого предка.
Наконец, можно столкнуться с адаптивным рядом, показывающим развитие какого-либо приспособления. Такой ряд может быть частью филогенетического ряда, например, приспособление ноги лошади к бегу, но часто этого нет, и адаптивный ряд можно составить даже за счет современных форм, вовсе не образующих филогенетического ряда. Как видим, палеонтологу приходится сталкиваться с большими трудностями. Его материал отрывочен, не полон.
Некоторым восполнением неполноты палеонтологических данных является, однако, возможность распространения на палеонтологию экологических данных. Определенная форма органа (строение ноги, структура зубного аппарата и пр.) позволяет делать заключения об образе жизни и даже о составе пищи вымерших животных. Отсюда возникает возможность реконструкции экологических отношений их. Заложенная в работах В. О. Ковалевского соответствующая область знания получила название палеобиологии (Абель, 1912). Она восполняет фрагментарность представлений палеонтолога о вымерших животных. В отношении форм, лишенных скелета, палеонтология дает лишь ничтожный материал по филогенетике, и в этих случаях на первое место выступает сравнительная морфология с ее методом сравнительного изучения гомологичных структур взрослых и зародышевых форм геологической современности. Отсутствие палеонтологических данных делает построение филогенетических выводов значительно более трудным. Поэтому наши филогенетические построения наиболее достоверны в отношении тех форм, для которых известен палеонтологический материал.
Все же исследователь и при отсутствии палеонтологических данных не остается безоружным. Он пользуется в этом случае другим методом, а именно, изучением стадий онтогенетического развития.