Отбор является одним из основных методов селекции.
В сочетании с генетическими методами он позволяет создавать новые формы, сорта и плоды с определенными свойствами и признаками. Наши выдающиеся советские селекционеры В. Я. Юрьев, А. П. Шехурдин, П. Н. Константинов, В. Н. Мамонтова, П. П. Лукьяненко, В. С. Пустовойт и многие другие создали ряд высокоурожайных сортов зерновых, масличных и технических культур. Так, например, В. С. Пустовойт, работая над повышением масличности семян Подсолнечника, в течение 15 лет увеличил ее с 29,44 (1940 г.) до 38,75% (1955 г.). Только благодаря созданию ценных сортов Производительность труда, выход масла на заводах, перерабатывающих эти семена, повысились более чем на 10%. В настоящее время масличность некоторых сортов подсолнечника еще более повышена. И эти сорта получили мировое признание.
Указанные успехи так же, как и весь мировой опыт селекции, достигнуты на основе применения различных методов отбора. Учение о методах отбора включает и различные приемы оценку признаков и свойств организмов, позволяющие наиболее точно определить их наследственные возможности.
В систему методов отбора входят два основных типа отбора: массовый и индивидуальный.
Массовый отбор
В самом элементарном представлении массовым называют отбор особей по внешним показателям (фенотипу), отвечающим принятому стандарту для популяции сорта или породы без проверки генотипа отдельных особей.
Так, при массовом отборе из всей популяции кур породы леггорн в хозяйствах оставляют для размножения матерей с яйценоскостью 150—200 яиц, живым весом 1,8 кг, не проявляющих инстинкта насиживания, т. д. Все куры, не отвечающие этим требованиям, выбраковываются из стада. При этом потомство каждой курицы и петуха индивидуально не оценивается, т. е. оценка производится исключительно по фенотипу. Фенотип является проявлением нормы реакции генотипа и в сильной степени обусловлен случайными коленями факторов внешней среды. В силу этих обстоятельств отбор по фенотипу оказывается недостаточно эффективным для оценки генотипов. Так, например, О. В. Гаркави показал, что средние удои коров, являющихся дочерями от лучших и худших по продуктивности матерей из одной популяции, не выходят за пределы средних удоев по стаду. Дочери от матерей с удоем 5529 ± 51,5 кг молока дали 4487 ± 108,8 кг, а дочери от матерей, имевших удой 3221,6 кг дали 3724 ± 80,9 кг. Средний же удой всех матерей данной популяции составлял 4115 ±24,8 кг, а средний удой дочерей 4375 ± 26,7 кг. Из этого мы видим, что массовый отбор по молочной продуктивности матерей в среднем для популяции оказывается малоэффективным.
Однако массовый отбор по некоторым другим признакам, как, например, жирность молока у коров или вес яиц кур, может давать некоторый сдвиг, поскольку коэффициент наследуемости этих признаков значительно выше.
Для иллюстрации зависимости эффекта массового отбора от коэффициента наследуемости отбираемого признака приведем следующий пример.
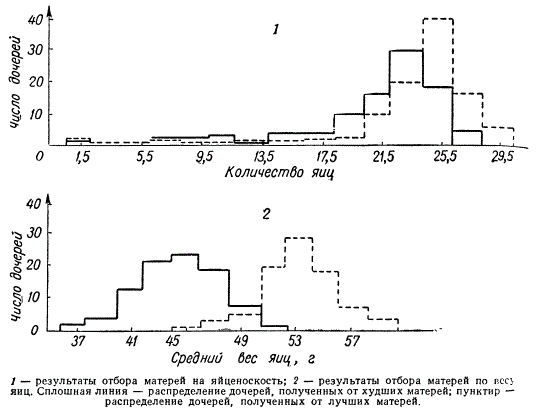
Эффективность массового отбора в зависимости от коэффициента наследуемости отбираемого признака (данные за месяц)
При коэффициенте наследуемости яйценоскости h2 = 0,25 дочери лучших и худших матерей в среднем снесли за один и тот же месяц приблизительно одинаковое количество яиц. Но дочери от матерей, откладывающих более крупные яйца, несли яйца более крупные, а от матерей, несших меньшие по размеру яйца, давали яйца более мелкие. Последнее объясняется тем, что коэффициент наследуемости веса яиц (h2) в этом случае равен 0,75. При таком высоком значении коэффициента массовый отбор оказывается эффективным в первом же поколении, хотя в последующих поколениях по мере повышения продуктивности он постепенно теряет свою эффективность. Таким образом, массовый отбор эффективен только для признаков с высокой наследуемостью.
Массовый отбор является медленно действующим средством улучшения популяции животных и растений, но он необходим и используется в определенных звеньях селекционной работы. Без его применения породы и сорта могут быстро утратить свои качества в процессе сельскохозяйственного производства.
Породы и сорта так называемой народной селекции (местные) создавались в течение длительного времени с помощью массового отбора. Такая местная порода, как романовская овца, многоплодная, с хорошими шубными качествами овчины, создавалась длительное время путем массового отбора в довольно ограниченном районе. То же можно сказать и об ярославском крупном рогатом скоте.
Итак, при массовом отборе вследствие отсутствия прямой оценки наследственных свойств родительского поколения успех селекции достигается медленно. Эффективность массового отбора зависит от коэффициента наследуемости признака, а также от размера популяции, в которой он осуществляется. Чем более гетерогенна популяция, внутри которой осуществляется массовый отбор, тем скорее может быть достигнут селекционный успех. Последний зависит также от того, насколько жестко проводится отбор. И далее, если популяция характеризуется низкой продуктивностью, но достаточно гетерогенна, то массовый отбор скорее дает эффект. При начальной высокой продуктивности исходной популяции массовый отбор оказывается малоэффективным.
Индивидуальный отбор
В отличие от массового отбора, при котором потомства разных организмов обезличиваются, при индивидуальном отборе, прежде всего, оценивается потомство отдельного растения или животного в ряду поколений.
Вследствие этого становится возможным оценивать наследственные качества отдельных индивидуумов: способность передавать свои свойства и норму реакции генотипа потомству.
В процессе индивидуального отбора популяцию искусственно разлагают на отдельные линии и семьи. При этом оценку и отбор животных по продуктивности производят по показателям всего или части потомства отдельной особи. В этой работе нередко применяют инбридинг, который и позволяет отбирать определенные генотипы, а также повышать уровень концентрации ценных генов в отдельной линии и, таким образом, увеличивать число гомозиготных особей в потомстве. Линии и семьи с лучшими показателями используют в дальнейшей селекционной работе. На племя оставляют таких особей, которые дают наибольшее число потомков с желаемыми свойствами, а остальных выбраковывают.
При индивидуальном отборе используют два метода. Одним из них является проверка производителя по потомству. Приведем пример. Были взяты две курицы — одна (№ 46) снесла за год 262 яйца, а другая (№ 12) за то же время — 258 яиц. Обе курицы скрещивались с одним и тем же петухом. Для проверки их наследственных свойств от каждой было взято по 7 дочерей без выбора. Продуктивность этих дочерей в одинаковых условиях была различной.

Проверка генетической обусловленности продуктивности кур по яйценоскости дочерей
Как видно из сравнения яйценоскости дочерей, курица № 12 более устойчиво передает высокую яйценоскость потомству, чем курица № 46. Если бы отбор матерей производился без учета яйценоскости потомства, а только по фенотипу, то казалось бы выгоднее оставить курицу № 46, так как она снесла несколько больше яиц. Однако после проверки по потомству становится ясно, что необходимо оставить на племя курицу № 12, имеющую более ценный генотип, так как она более надежно передает свойства яйценоскости.
При индивидуальном отборе на продуктивность необходимо вести оценку как самок, так и самцов. Хотя яйценоскость и молочность проявляются только у женского пола, генотип мужского пола играет очень важную роль в наследственном определении этих признаков. Например, петухи породы род-айланд имеют различную племенную ценность, определяемую различием генотипов. Наиболее ценным по генотипу является петух № 173.

Различная генетическая ценность петухов-производителей, определяемая по яйценоскости дочерей
Оценка и отбор производителей по потомству представляют наиболее надежный путь для совершенствования породных качеств. Именно поэтому принцип индивидуального отбора, в частности метод испытания по потомству, нашел широкое применение в селекции сельскохозяйственных животных.
Другим методом индивидуального отбора является метод сиб-селекции. Как мы видели, генетическая ценность петухов (способность к передаче свойства высокой яйценоскости) может быть определена по дочерям, наследственные качества быка — по молочности его дочерей и т. д. Наследственные свойства можно оценивать по продуктивности родственных особей — братьев или сестер. Сиб-селекция и означает оценку и отбор по братьям и сестрам (sibling по-английски «брат-сестра»). Примером сиб-селекции может служить отбор генетически ценных петушков по продуктивности (яйценоскости) их родных сестер. Если сестры имеют высокую яйценоскость, то есть основание считать, что и братья обладают таким генотипом, который может обеспечить высокую яйценоскость их дочерей.
Другим примером сиб-селекции может служить отбор свинок по откормочным качествам на основании контрольного откорма хрячков из того же помета. В этом случае наследственно определяемые откормочное качества хрячков, т. е. скороспелость, оплата корма, развитие отдельных частей тела, связанных с мясностью, дают основание оценить по этим же показателям и поросят-свинок. В Дании принято из одного помета брать на контрольный откорм да два поросенка того и другого пола. В случае хороших результатов остальная часть приплода оставляется на племя. В случае плохих результатов весь помет выбраковывается. Таким способом датским селекционерам удалось за 10 лет снизить затрату кормов на 1 кг привеса с 6,48 до 5,52 кормовых единиц. В целом по стране это дало большой экономический эффект. В последние годы разработана методика определения откормочных качеств свиней по оплате корма по измерению толщины сала на живых особях.
Рассмотрим пример сиб-селекции. Для того чтобы вывести линию мух или других насекомых, устойчивых к яду, необходимо воздействовать на них этим ядом. Обычно после такой обработки отбирают выживших особей и получают от них потомство. Таким путем из поколения в поколение с помощью отбора удается повышать резистентность к яду. При указанном методе отбора трудно доказать, что отбор генотипов явился причиной возникновения наследственно обусловленной резистентности, так как можно думать, то в этом случае произошла онтогенетическая адаптация. С помощью сиб-селекции это сомнение легко разрешить. Потомство жаждой отдельной пары мух в каждом поколении разделяют на две группы. Одну половину мух подвергают обработке ядом, а других — оставляют без воздействия. Если обработанные ядом особи из данной семьи обнаруживают высокую резистентность, то от их братьев и сестер, не соприкасавшихся с ядом, получают потомство. Исследующем поколении повторяют ту же процедуру обработки и отбора. И так из поколения в поколение на основании сиб-селекции осуществляют отбор насекомых, не соприкасавшихся с ядом.
В результате удается получать высокорезистентные линии. Так, Ю. М. Оленову и другим исследователям с помощью сиб-селекции удалось во много раз повысить резистентность дрозофилы к ДДТ и другим ядам.
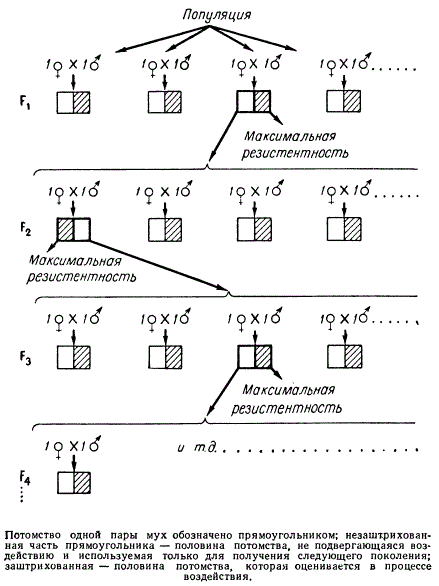
Схема сиб-селекции
Подобная методика давно применяется в селекции растений, где этот прием называется методом половинок. Применялся он и для изучения устойчивости микроорганизмов к антибиотикам.
Результаты сиб-селекции доказывают, что само воздействие факторами служит лишь индикатором, вскрывающим генетическую потенцию линии. Действие внешнего фактора — яда в приведенном примере — никак не затрагивает отбираемые организмы, и тем не менее в ряду поколений происходит повышение резистентности, что и указывает на отбор генотипов.
Индивидуальный отбор является основным методом в селекции животных, растений и микроорганизмов. Его применение особенно облегчается у растений-самоопылителей, где потомство одной особи составляет чистую линию. У перекрестноопылителей индивидуальный отбор производится так же, как и у животных, — по оценке семей и линий.
Итак, индивидуальный отбор является наиболее верным средством для оценки и создания определенных генотипов в процессе селекции. При этом надо иметь в виду, что порода и сорт создаются для определенных условий, в которых они будут существовать. И поэтому нельзя ожидать от одной и той же породы или сорта в разных условиях тождества продуктивности. Несмотря на то, что отбор в конечном счете оценивает генотип организма, его действие зависит от внешней среды. Отбор, осуществляющийся на фоне среды, максимально выявляющей наследственные возможности (норму реакции генотипа) отбираемых организмов, более эффективен, чем в других условиях. Очевидно, что нельзя вести отбор на засухоустойчивость растений во влажных климатических условиях, на морозоустойчивость — при высоких положительных температурах, на длинный световой день — в условиях короткого дня, на иммунность растения — в условиях отсутствия заражения и т. д.
Соответствующие условия внешней среды облегчают оценку генотипа, делают ее наиболее объективной и точной. Именно в целях более полной оценки генотипа часто требуется создание крайнего или наиболее оптимального фона внешней среды, наиболее точно выявляющего отбираемые генотипы. Испытание генотипа в крайних условиях иногда называют отбором на провокационном, или селективном, фоне.
В заключение необходимо сделать несколько общих замечаний по поводу понимания действия факторов среды и роли отбора в эволюционном процессе и в селекции.
1. На основании огромного опыта генетики пришли к выводу, что нарушение равновесия между организмом и внешней средой, изменяя систему обмена веществ, вызывает наследственную изменчивость. Мутации могут быть как вредными, так и полезными, но в процессе эволюции в результате отбора сохраняются и накапливаются такие, которые обеспечивают адекватное приспособление к среде.
2. Характер и темп отбора тесно связаны как с мутационной, так и с комбинативной изменчивостью. При неблагоприятных условиях внешней среды происходит изменение направления и темпа отбора и одновременно направления и темпа изменчивости.
3. Наследственные изменения типа мутаций не адекватны факторам, определяющим направление отбора, но они могут быть адекватны факторам, вызывающим мутации.
4. Факторы внешней среды, играющие селекционную роль, действуют двояко: в генотипе — на изменение гена, в фенотипе — на действие гена. Поэтому, признавая мутации как основу наследственной изменчивости, нельзя говорить об их адекватности изменениям факторов среды.
5. Процесс эволюции организмов адекватен внешней среде: рыбы приспособлены к водной жизни, млекопитающие к наземной, и т. д.; каждый вид «пригнан» к определенным условиям жизни. Это соответствие со средой создается естественным отбором путем переживания наиболее приспособленных организмов. Эволюция домашних животных и возделываемых растений идет в соответствии с условиями их существования и создается искусственным отбором.
6. С помощью различных методов отбора на основе комбинатовной и мутационной изменчивости генетика в комплексе с другими дисциплинами современного естествознания может конструировать новые формы жизни.
Изучение генетических основ селекции позволило подвести научную базу под эмпирические приемы селекции растений животных и микроорганизмов, т. е. объяснить значение различных традиционных методов отбора и методов скрещивания.
Генетика разработала принципиально новые подходы к селекции, ускоряющие темпы создания новых форм, а именно:
- искусственное вызывание наследственной изменчивости с помощью ионизирующей радиации и химических мутагенов;
- создание межлинейных гибридов различной сложности для практического использования явления гетерозиса как у растений, так и у животных;
- применение цитоплазматической мужской стерильности, открывшей путь производства межлинейных гибридных семян перекрестноопылителей (кукуруза, сорго и др.) и самоопылителей (пшеница);
- применение полиплоидии для повышения продуктивности сельскохозяйственных растений;
- получение биохимических мутаций и отбор их на селективных средах, что открыло новые принципы промышленного производства аминокислот, витаминов, антибиотиков и других органических веществ.
Несмотря на очевидные успехи генетики, в ней остается много неизведанного для использования резервов природы на благо человека. Главным условием прогресса в этом направлении является глубокое изучение генетики отдельных видов растений, животных и микроорганизмов, т. е. частной генетики.