Функциональный тест на аллелизм основан на том, что зигота, несущая две мутации одного гена в транс-положении, имеет мутантный фенотип.
В 1956—1957 гг. при исследовании мутантов нейроспоры оказалось, что некоторые аллели одного гена способны при объединении в гетерокарионе давать начало культуре дикого или близкого к дикому типа. Так, если получать гетерокарионы между мутантами, нуждающимися в аденине вследствие мутации в локусе, контролирующем фермент аденилосукциназа, то некоторые пары мутантов образуют гетерокарионы, способные расти на среде без аденина. Аналогичные случай так называемой комплементации аллелей были обнаружены вскоре для целого ряда генов как у нейроспоры, так и у других генетических объектов от микроорганизмов до дрозофилы.
Особенностью такой межаллельной комплементации было то, что лишь некоторые строго постоянные сочетания аллелей могли образовать зиготу дикого или близкого к нему типа. Эту закономерность во взаимодействии аллелей можно изобразить графически, т.е. построить карту межаллельной комплементации. Каждую аллель, точнее мутацию каждой аллели, принято изображать в виде отрезка прямой. Если две аллели при объединении их в зиготе комплементируют, т. е. восстанавливают в зиготе дикий фенотип, то соответствующие отрезки изображают так, чтобы они не перекрывали друг друга. Если же аллели не комплементируют, то отрезки изображают перекрывающими друг друга. Как правило, для каждого гена можно построить лишь одну карту комплементации.
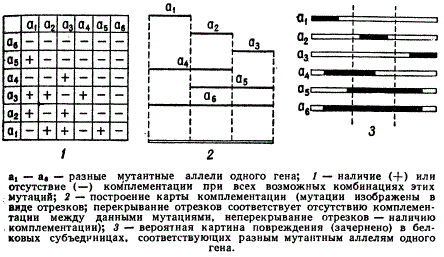
Межаллельная комплементация
Аллели а1, а2 и а3 комплементируют между собой во всех сочетаниях. Соответствующие отрезки должны быть изображены неперекрывающимися. Порядок их изображения однозначно определяется характером комплементирования аллелей с мутациями a4, a5. Поскольку а4 комплементирует только с а3, то следует а3 изобразить справа от а1 и а2. Аналогично, характер комплементирования а5 требует изображения а1 слева от а2 и а3. При испытании большого числа аллелей (десятки и сотни) одного гена обнаруживается, что некоторые из них комплементируют одинаково, например, по типу а1 или а4 или а5 и т. д., т. е. аллели по характеру комплементирования могут быть разбиты на группы: группа а1, группа а2 и т.д., в пределах которых комплементации не происходит. Поэтому отрезки различной длины, образующиеся при составлении карт комплементации, называют группами комплементации. Если эти отрезки спроектировать на одну прямую, то получаются так называемые единицы комплементации, или комплоны.
Число групп так же, как и число единиц комплементации, является отражением того, сколько аллелей испытано в опытах по изучению аллельной комплементации данного гена. Масштаб изображения групп комплементации обычно произволен, однако группы комплементации изображаются таким образом, чтобы при их проецировании образовались комплоны равной длины. Мутации тех аллелей, которые вообще не способны комплементировать, изображаются напрерывной линией, перекрывающей все комплоны данного гена, например группа а6 на рисунке.
Исследование механизма межаллельной комплементации произвел Д. Вудвард, который в 1959 г. показал, что эффект комплементирования аллелей можно воспроизводить in vitro на бесклеточных экстрактах мутантов по аденилосукциназе у нейроспоры. Активность этого фермента в экстрактах двух мутантов, способных к межаллельной комплементации, была равна нулю. Если же экстракты тех же самых мутантов смешать, то появляется ферментативная активность, которая, правда, никогда не достигает 100%, свойственных экстрактам из штамма дикого типа, и даже 50 %, которых следовало бы ожидать для гетерозигот или гетерокарионов.
Таким образом, было показано, что механизм межаллельной комплементации заключается во взаимодействии генных продуктов (белковых молекул или субъединиц) в цитоплазме. Низкая ферментативная активность при этом указывает на отличие комплементирования аллелей от комплементирования мутантных генов.
Дальнейшее исследование явления межаллельной комплементации показало, что существует строгая корреляция между обнаружением самого явления в тех или иных локусах и строением белка, который контролирует исследуемый ген. Известно, что существуют ферменты, которые способны осуществлять свои функции только после образования так называемой четвертичной структуры, т. е. только после того, как несколько одинаковых белковых глобул — фактически копий одного гена — объединяются между собой. Принято говорить, что такие ферменты построены из идентичных субъединиц.
Межаллельная комплементация была обнаружена до сих пор только для тех генов, которые контролируют ферменты, состоящие из идентичных субъединиц. Это наводит на мысль о возможном взаимодействии идентичных субъединиц, несущих различные мутационные изменения, в результате которого происходит как бы взаимное исправление мутационных дефектов в целой молекуле фермента. Это предположение было подтверждено в 1965 г. работами Дж. Финчема и А. Коддингтона, которые смешивали очищенные мутантные белки (лишенные ферментативной активности), синтезируемые под контролем различных аллелей гена am у нейроспоры.
Ген am контролирует фермент глютомат-дегидразу, который построен из идентичных субъединиц. Используя радиоактивную метку, Дж. Финчем и А. Коддингтон показали, что ферментативная активность в смеси мутантных белков проявляется только при объединении по-разному мутантных субъединиц этого фермента.
Правда, до сих пор остается открытым вопрос о природе взаимодействия, приводящего к взаимному исправлению мутационных Повреждений, которые несет каждая из субъединиц. Предполагается, что всякая аллель, способная к межаллельной комплементации, детерминирует синтез белковой молекулы (субъединицы) с большим или меньшим по протяженности изменением в структуре молекулы.
Предполагается, что даже если в гене изменена всего одна пара нуклеотидов (точковая мутация), а в полипептиде соответственно заменена лишь одна аминокислота по сравнению с полипептидом дикого типа, то это может привести к протяженному изменению молекулы белка, к изменению конформации (складывания) молекулы. При этом комплементировать будут только те субъединицы (фактически те аллели), которые несут не перекрывающиеся, де заходящие при объединении субъединиц изменения, что выражается на карте комплементации в виде неперекрывающихся отрезков (групп комплементации).
Таким образом, хотя в межаллельной комплементации еще не ясны некоторые моменты, уже сейчас это явление служит мощным оружием генетиков в изучении структуры и функции индивидуальных белков — ферментов in vivo.
Карты комплементации обычно имеют довольно сложную структуру. Примером может служить карта комплементации аллелей локуса ad2, контролирующего один из этапов в синтезе аденина и дрожжей Saccharomyces cerevisiae, построенная на основании гетероаллельных скрещиваний, проведенных Т. Сойдла.

Предварительная карта комплементации локуса ad2y дрожжей Saccharomyces cerevisiae
На карту нанесены два типа аллелей: те мутантные аллели, присутствие которых в клетке приводит к полному выключению синтеза аденина, и те аллели, которые контролируют синтез аденина, но очень в слабой степени. Очевидно, что мутанты двух типов должны иметь соответственно различные повреждения в молекулах фермента, контролируемого геном ad2. Видимо, фермент мутантов, у которых синтез аденина блокирован полностью, должен иметь и более ощутимые повреждения в структуре молекулы, нежели фермент мутантов лишь с заторможенным синтезом аденина. Действительно, что аллели первого типа несут изменения, захватывающие от одного комплона до 20, т. е. фактически среди них имеется значительный процент аллелей, вовсе не способных Комплементировать. В то же время аллели второго типа несут Изменения, распространяющиеся не более чем на 5 комплонов. Его доказывает, что анализ карт комплементации позволяет судить о характере изменения белка, контролируемого данным геном. Изучая межаллельную комплементацию, в частности при построении карты, удалось установить влияние целого ряда внешних агентов на этот процесс. Например, обнаружено влияние концентрации ионов калия и магния в среде на способность некоторых аллелей комплементировать. Показано также существование генов-модификаторов межаллельной комплементации и влияние генов-супрессоров на характер комплементирования аллелей.
Несмотря на все сказанное, функциональный критерий аллелизма может дать ответ о принадлежности рассматриваемых мутаций к одному гену. В случае генов, контролирующих ферменты, не имеющие субъединичного строения, ответ будет однозначном при испытании любой пары мутантов. В случае генов, контролирующих ферменты, построенные из идентичных субъединиц, необходимо иметь серию мутантов, при скрещивании которых комплементирование аллелей выразится в сложных взаимоотношениях, что позволит построить карту комплементации. Все аллели исследуемого гена будут комплементарны любой аллели другого гена. Среди аллелей каждого гена существует группа вообще не комплементирующих. Использование таких аллелей для функционального теста позволяет определить принадлежность испытуемой мутации к данному гену.