Наряду с непосредственным участием минеральных элементов в поддержании гомеостаза и обменных процессах возможно их косвенное влияние на метаболизм через симбиотическую микрофлору.
Относительно постоянные и благоприятные условия способствуют интенсивной деятельности микрофлоры в разных отделах пищеварительного тракта — в преджелудках и слепой кишке у жвачных, в слепой и ободочной кишках у лошадей, в слепой кишке у свиней и кроликов, в зобе, мышечном желудке и слепых отростках у птиц. Можно утверждать, что нормальное осуществление пищеварительной функции у животных невозможно без деятельности микрофлоры. Особенно это касается жвачных, преджелудки которых населены многочисленной и разнообразной по составу микрофлорой и микрофауной, принимающими активное участие в переваривании корма.
Априори можно утверждать, что микрофлора пищеварительного тракта нуждается в тех же группах питательных веществ (включая минеральные элементы), что и макроорганизм, хотя пути их превращения у симбионтов могут быть принципиально иными.
О важной роли минеральных элементов для деятельности микрофлоры свидетельствуют:
- увеличение количества и активности отдельных групп микроорганизмов при введении в рацион солей макро- и микроэлементов (в смеси или в отдельности);
- положительное влияние добавок золы люцерны или меляссы на переваримость жвачными целлюлозы низкокачественных грубых кормов;
- усиление биосинтеза микробного белка и снижение потерь азота при восполнении дефицита микроэлементов;
- изменение метаболизма (всасывания и экскреции) минеральных веществ у моногастричных под влиянием ряда антибиотиков, изменяющих соотношение популяций микроорганизмов;
- более низкая потребность в некоторых минеральных веществах (Mg, Zn, Ca, Р) и более высокая их абсорбция у стерильных животных (в частности, цыплят) в сравнении с конвенциональными (т. е. зараженными кишечной микрофлорой);
- резкое уменьшение содержания водорастворимых фракций микроэлементов в рубцовом содержимом в сравнении с кормом и химусом двенадцатиперстной кишки (связывание микроэлементов микрофлорой).
Очевидно, потребность микрофлоры в макро- и микроэлементах может зависеть от состава рациона. Так, при зерновом рационе быстрорастущие микроорганизмы имеют повышенную потребность в минеральных веществах в сравнении с медленнорастущими, переваривающими в основном грубые корма. При скармливании бобовых культур (богатых микроэлементами) потребность микрофлоры ниже, чем при использовании злаковых культур.
Потребность рубцовых микроорганизмов в минеральных элементах определяли in vitro с использованием разных вариантов методики «искусственного рубца». Субстратом служила рубцовая жидкость с добавками или без добавок синтетических источников азота и клетчатки и с разными дозами изучаемых элементов. Критерием эффективности добавок служили: рост числа бактерий, переваривание целлюлозы, образование ЛЖК, уменьшение содержания небелкового азота и увеличение количества азота микроорганизмов.
В этих опытах было установлено, что рубцовые бактерии (иногда их отдельные виды) нуждаются в Ca, P, Na, К, Mg, Cl, S, Mn, Cu, Co, Fe, Mo, Zn.
При культивировании отдельных видов микроорганизмов на питательных средах в состав последних также вводят минеральную смесь.
По аналогии этот перечень можно распространить и на кишечную микрофлору. Сведения, полученные in vitro, в целом согласуются с опытными in vivo. Простейшие в сравнении с бактериями менее активно реагировали на изучаемые добавки in vitro, что предполагает их большую потребность в микроэлементах.
Примерные оптимальные и токсические концентрации макро- и микроэлементов для микрофлоры приведены в таблицах.
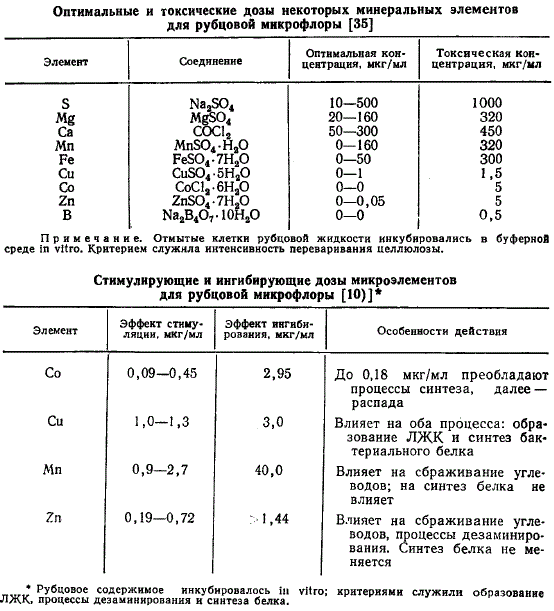
Приведенные в таблицах результаты в целом сравнимы. Вместе с тем они подтверждают точку зрения, что эффект минеральных добавок зависит от условий опыта: выбранного критерия, субстрата, источников азота и углеводов, времени инкубации. Так, при наличии легкосбраживаемых углеводов в среде концентрация микроэлементов может оказаться более высокой без токсического эффекта.
При культивировании разных видов рубцовых бактерий на агаровой среде оптимальная потребность их в натрии определена в 40—400 мг/л. Ионы рубидия, цезия и лития в тех же дозах оказались токсичными. Калий же почти во всех случаях мог быть заменен рубидием.
Имеются сведения о положительном влиянии на целлюлозолитическую микрофлору рубца добавок йода.
Анализ литературных данных позволяет предположить существование следующих механизов взаимодействия минеральных элементов с симбиотической микрофлорой желудочно-кишечного канала.
1. Элемент необходим и макроорганизму, и микроорганизмам, т. е. имеет место его конкурентное потребление. Требующиеся для макроорганизма вещества доставляются ему при переваривании микробных тел.
К числу таких элементов относятся: кальций (необходимый для роста некоторых видов микроорганизмов, например b. succinogenos), фосфор (необходимый для энергетических процессов и размножения клеток), магний, железо, цинк, молибден (как составные части или активаторы бактериальных ферментов).
2. Элемент необходим только (или главным образом) микроорганизмам; последние же продуцируют метаболиты, нужные макроорганизму.
Примером может служить кобальт, соли которого являются по существу питательным материалом для микрофлоры. Основная роль кобальта — обеспечение потребностей некоторых групп бактерий, продуцирующих необходимый животным витамин B12 или его аналоги (кобамины, кобаламины). Это происходит в преджелудках жвачных, толстом кишечнике моногастричных животных, в слепых кишках кроликов и кур.
3. Элемент необходим обоим симбионтам, но в данной форме может быть ассимилирован лишь микроорганизмами.
В качестве примера можно назвать неорганическую серу, которая необходима для переваривания целлюлозы, усвоения небелкового азота, синтеза витаминов группы В.
Млекопитающие, как известно, лишены ферментов, обеспечивающих использование сульфидной серы. Образующийся H2S окисляется у них до серной кислоты, которая затем нейтрализуется. Потребность в ионах SO42--удовлетворяется, таким образом, за счет сульфатов, образующихся при катализе серосодержащих аминокислот.
Бактерии способны восстанавливать сульфаты до сульфидов, а последние включать в серосодержащие аминокислоты и белки. Аминокислоты в рубце, следовательно, могут синтезироваться с использованием углеводов, аммиака и минеральной серы.
Аналогичный процесс наблюдается и в толстом кишечнике моногастричных животных. У птиц, наряду с этим в процессе межуточного обмена происходит в небольшой мере синтез из сульфата таурина с последующим образованием цистеина.
4. Элемент необходим в обменных процессах макроорганизма, но одновременно он участвует в создании оптимальной среды для жизнедеятельности микроорганизмов.
Представителями таких элементов являются калий, натрий, хлор, отчасти фосфор.
Сложный желудок у жвачных животных представляет собой замкнутую биологическую систему, поддерживающую постоянство своей внутренней среды (буферность, осмотическое давление, соотношение ионов) за счет поступлений извне, выделения слюны, двусторонней проницаемости стенки преджелудков. Особое значение при этом имеют катионы К+, Na+, NH4+, анионы Cl, НСО3—, НРО42-. Ионы Na+ и Cl— проникают через стенку в кровь путем активного транспорта против градиента концентрации и электрохимического градиента (хлориды переходят лишь при низкой концентрации К+ и высокой Na+). Ионы К+, NH4+ и НРО42- всасываются путем диффузии, причем фосфатные ионы в незначительном объеме. Ионы НСО3— активно транспортируются в кровь; переход их в обратном направлении происходит в порядке обмена на ЛЖК.
Поддержание оптимальной среды является функцией макроорганизма, и в этом большая роль принадлежит рубцовой стенке, избирательно проницаемой в обоих направлениях. При недостатке в рационе отдельных минеральных элементов наблюдается тенденция выравнивания среды за счет ресурсов макроорганизмов. Это доказано опытами на животных с изолированным участком рубца. В самой рубцовой стенке могут создаваться запасы макро- и микроэлементов, которые вступают в ковалентные связи с белками и фосфолипидами.
Рассмотренные основные функции минеральных элементов в организме, по-видимому, не исчерпывают всего многообразия их возможного влияния на метаболические процессы. В литературе есть сведения о влиянии макро- и микроэлементов на функцию пищеварительных желез, абсорбцию питательных веществ, процессы кроветворения, тонус сосудов, специфический и неспецифический иммунитет и др. Однако все эти функции являются, по-видимому, лишь конкретным проявлением описанных выше основных биохимических механизмов.
Снижение же продуктивности и нарушение воспроизводительной функции у животных при недостаточном или несбалансированном минеральном питании, за некоторым исключением, нужно рассматривать как следствие общего расстройства обмена веществ и ослабления резистентности организма.