До сих пор при изложении материала мы исходили из положения, что один и тот же локус гомологичных хромосом представлен двумя аллелями: A и a, В и b, С и с и т. д.
Эти два состояния локуса возникают при прямом и обратном мутировании. На самом деле один и тот же ген может изменяться в несколько состояний; иногда таких состояний бывает несколько десятков и даже сотен. Ген А может мутировать в состояние а1, а2, а3, …, аn или ген В в другом локусе — в состояние b1, b2, b3, …, bn и т. д. Мутации одного и того же локуса называют серией множественных аллелей, а само явление — множественным аллелизмом.
Возникновение серии множественных аллелей схематически иллюстрируется на рисунке.
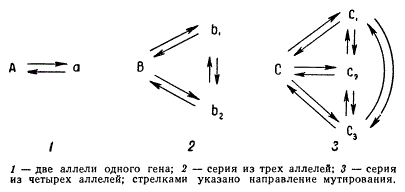
Схема возникновения серии множественных аллелей
Изучение мутаций серии множественных аллелей показало, что:
- любая аллель такой серии может возникать мутационно непосредственно от аллели дикого типа или любого другого члена данной серии;
- любая аллель серии может мутировать в другую как в прямом, так и в обратном направлении;
- каждый из членов серии, по-видимому, имеет свою характерную частоту мутирования;
- серии множественных аллелей в разных локусах могут иметь различное число членов.
Наследование членов серии множественных аллелей подчиняется менделевским закономерностям. При этом имеет место следующее:
1) серия множественных аллелей у каждого диплоидного организма может быть представлена одновременно только двумя любыми ее членами, например:
Аа1, Аа2, а1а2, а1а3, а2а2 и т. д.;
2) каждый из членов серии может полностью или не полностью доминировать над другим ее членом, например: A > a1 > a2 > a3 и т.д.
3) члены одной серии действуют на один и тот же признак; одновременно они могут иметь множественный эффект.
Рассмотрим более подробно наследование серии аллелей одного гена, а также приведем факты множественного аллелизма.
У грызунов, в частности у кроликов, существует серия множественных аллелей по окраске шерсти: черный, шиншилла, гималайский горностаевый; — неполный альбинос и полный альбинос (белый кролик с красными глазами). При скрещивании черных кроликов с гималайскими, имеющими на фоне общей белой окраски шерсти черные кончики ушей, лап, хвоста и морды, в F1 все потомство оказывается черным. Во втором поколении наблюдается расщепление в отношении 3 черных к 1 гималайскому. Скрещивание гималайского кролика с альбиносом дает гибридов F1 с признаками первого родителя, а в F2 имеется расщепление 3 гималайских на 1 альбиноса.

Схема наследования серии множественных аллелей
Следовательно, каждая пара членов данной серии ведет себя в расщеплении как одна аллельная пара. Если бы члены серии были неаллельными, то должно было происходить расщепление, соответствующее дигибридному или полигибридному скрещиванию. Однако этого не наблюдается. При проверке других мутаций в этой серии во всех случаях имеет место моногибридное расщепление.
На основании подобного генетического анализа предположили, что ген альбинизма может иметь несколько состояний. Обычно такую серию обозначают по названию признака, впервые найденного, или по общему характеру действия данного локуса, способного мутировать в разные состояния. Так, например, серия множественных аллелей гималайского альбинизма у кролика обозначается: буквой С — черный, а члены ряда в гомозиготном состоянии cch cch — шиншилловая окраска, chch — гималайский альбинос, саса — полный альбинос.
В отличие от генов, для которых известны только два состояния, сочетание двух разных членов серии множественных аллелей в гетерозиготе называют компаундом.
В связи с тем, что у диплоидного организма могут присутствовать одновременно только два члена серии множественных аллелей, представляет интерес выяснить, как они ведут себя в разных сочетаниях. Члены ряда серии аллелей не только по-разному определяют развитие признаков, но и вступают в разные доминантно-рецессивные отношения друг с другом, о чем было упомянуто выше. Нередко доминирование при этом неполное. Так, например, компаунд по шиншилловой окраске и гималайской или по шиншилловой и альбинизму cchch или cchca дает светло-серую окраску, типичную для шиншиллы, а компаунд с са —фенотип гималайского кролика.
Альбинизм оказывается рецессивным по отношению ко всем членам данной серии. Аллель альбинизма по ранее приведенной классификации Мёллера является аморфной мутацией, а аллель гималайской окраски — гипоморфной мутацией.
Градуальность (промежуточность) проявления членов серий множественных аллелей в компаунде наблюдается в тех случаях, когда мутантные гены относятся к разным типам действия: аморфный — гипоморфный и т. д. У морской сцинки, например, так же, как и у кролика, имеется серия множественных аллелей по окраске шерсти. Окраска шерсти оказывается различной в зависимости от сочетания членов данной серии. С. Райт и его сотрудники изучили у морской свинки количество основного пигмента — меланина, обусловливающего у этого животного окраску шерсти. Серия аллелей окраски состоит из гена С и его мутантов: ck, cd, сr и сa.
Аллель С доминирует над всеми, а са, обусловливающая альбинизм, является рецессивной по отношению ко всем остальным членам данной серии, которые определяют промежуточную пигментацию шерсти. На основании колориметрического исследования экстрактов меланина из шерсти морских свинок разных компаундов данной серии удалось установить градуальное действие мутаций.
Число мутантных аллелей данного гена может быть весьма большим. Так, например, по гену белой окраски глаза (white) у дрозофилы известна серия аллелей из 12 членов. Все эти аллели в компаунде в определенной последовательности дают промежуточную окраску глаз и доминируют над аморфным геном w. Ниже приводятся сведения о фенотипических проявлениях аллелей (цвет глаз) этой серии и их символы:

У одного и того же вида растений или животных целый ряд локусов может быть представлен серией множественных аллелей. Такой ряд установлен у крупного рогатого скота, кроликов, мышей, морских свинок, дрозофилы, а также у кукурузы, табака, гороха и др. Серии множественных аллелей обнаружены и у человека. Распространенность этого явления среди животных, растений и микроорганизмов могла быть обусловлена несколькими причинами: множественный аллелизм увеличивает резерв мутационной изменчивости в эволюции, в силу чего он приобрел приспособительное значение.
Изучение множественного аллелизма имеет не только теоретическое значение для понимания природы наследственной изменчивости, но и чисто практическое. Так, у человека известны четыре группы крови: А, В, АВ и 0. Если взять кровь от человека группы АВ или А или В и перелить другому человеку, имеющему кровь группы 0, то последний может погибнуть. Причина этого заключается в следующем. Эритроциты группы АВ содержат два антигена: группа А — антиген А, группа В — антиген В, группа 0 не содержит антигенов А и В. Сыворотка крови этих четырех групп различается следующим образом: группа 0 имеет два антитела, обозначаемые как α и β; группа А содержит в сыворотке антитело β, группа В — антитело α, сыворотка группы АВ не имеет антител α и β. Взаимоотношение антигенов эритроцитов (А, В) антител сыворотки (α и β) всех четырех групп крови приведен в таблице.

Реакция агглютинации эритроцитов между различными группами крови
В ряде случаев группы крови оказываются несовместимыми. Происходит это потому, что антитело α агглютинирует эритроциты групп крови А и АВ, а антитело β — эритроциты групп крови В и АВ. Если в крови реципиента с группой А окажется антиген В, то наступит слипание эритроцитов донора; то же происходит, если в кровь реципиента группы В попадают антигены донора А или АВ.
Группы крови не изменяются в течение жизни человека. Причем их наследование осуществляется настолько четко, что это используется в судебной медицине для исключения отцовства и в некоторых других случаях. Для примера приведем ожидаемые и невозможные группы крови у потомков при различном сочетании групп крови родителей.
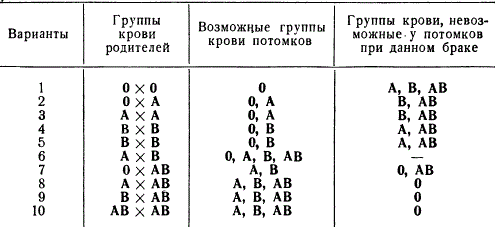
Группы крови потомков от браков людей с разными группами крови
Как показал А. Винер, в наследственном определении антигенов участвует серия множественных аллелей, состоящая по меньшей мере из трех членов: IА, IB, I0. Первые два члена серии определяют выработку антигенов, соответственно IА — антигена А, I0 — антигена В. Ни один из них не доминирует над другими. До недавнего времени считали, что I0 является рецессивной аллелью к первым двум аллелям, поэтому ее обозначают также i. Сочетание всех этих аллелей определяет соответствующие группы крови, а именно:

Гетерозиготы IАI0 и IBI0 не отличаются по фенотипу от гомозигот. В настоящее время генетические исследования групп крови, т. е. установление генов, определяющих антигенные различия, показывают, что каждая группа зависит от целого ряда аллелей однозначного действия (А1, А2, А3 или В1, В2, В3 и т. д.). Кроме того, некоторые авторы считают, что существуют люди с генотипами I0I0 (т. е. ii), имеющие 0 группу крови, эритроциты которых обладают антигенными свойствами и имеют соответствующие антитела.
Группы крови у крупного рогатого скота также определяются сериями множественных аллелей. Изучение наследственной детерминации групп крови составляет предмет исследования новой, чрезвычайно перспективной области генетики, называемой иммуногенетикой.
В настоящее время не совсем ясно, все ли локусы могут иметь серии множественных аллелей. Предполагалось, что последние обнаруживаются лишь для некоторых локусов хромосом. Но по мере исследования отдельных генов у наиболее изученных форм, в свете современных данных о строении гена, складывается впечатление, каждый локус может быть представлен серией множественных аллелей с большим или меньшим числом членов. Следует отметить, что у близких видов встречаются сходные серии аллелей (например, в пределах отряда грызунов и др.). Это говорит о гомологии наследственной изменчивости идентичных локусов хромосом у родственных видов.
Таким образом, исследование множественного аллелизма показывает, что ген как наследственная единица может мутировать в ряд состояний.