Голова
Тело кальмаров разделяется на голову и туловище. Голова отделяется от туловища ясно выраженным шейным перехватом, или шеей. На боковой поверхности шеи у многих видов есть продольные складки, разделенные гребнями. Обычно этих складок три. У некоторых видов, например, из семейства Onychoteuthidae имеются, кроме того, затылочные складки — дополнительные продольные гребни, образующие направляющие желобки для воды. По-видимому, благодаря им вода входит в мантийную полость не в виде сплошного кольца, повторяя форму мантийной щели, а в виде отдельных струй. Интересно, что дополнительные продольные гребни отмечены у ряда быстроплавающнх некто иных видов (Onychoteuthis banksi, Ancistroteuthis lichtensteini).
По бокам головы расположены большие округлые глаза, по высоте организации не уступающие глазам высших позвоночных животных. О важном значении глаз как органа чувств свидетельствует не только их сложное строение (наличие роговицы, радужины, хрусталика, стекловидного тела и сетчатки), но и значительные абсолютные и относительные размеры их. Диаметр глаза у некоторых гигантских кальмаров рода Architeuthis достигает 30—40 см. Относительная величина глаза у взрослых пелагических кальмаров составляет около 6—10% абсолютной длины тела. В онтогенезе величина и форма глаза не остаются постоянными: глаз несколько вытягивается вдоль продольной оси тела кальмара, а его относительные размеры уменьшаются. Особенно велики (по отношению к размерам тела) глаза личинок кальмаров.
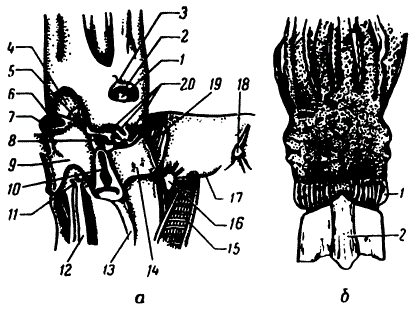
Голова и замыкательный аппарат кальмара: a — Todarodes pacificus, вид слева и немного снизу, мантия разрезана по брюху и отогнута: 1 — зрачок, 2 — радужная оболочка, 3 — вырезка (синус) века, 4 — вороночная ямка, 5 — мускул-подниматель (аддуктор) воронки, 6 — вороночный клапан, 7 — отверстие воронки, 8 — мускул-сжиматель воронки, 9 — воронка, 10 — вороночный хрящ, 11 — заднепроходное отверстие, 12 — прямая кишка, 13 — мускул-втягиватель воронки, 14 — шейный мускул, 15 — жабра, 16 — звездчатый мантийный ганглий, 17 — перепонка, соединяющая жабру с мантией, 18 — мантийный хрящ, 19 — затылочный хрящ, 20 — продольные шейные складки (по Иванову и Стрелкову, 1949); б — Onychoteuthis banksi, голова сверху, мантия удалена: 1 — затылочные складки, 2 — затылочный хрящ (по Muus,1959)
У относительно глубоководных видов абсолютные размеры глаза по сравнению с пелагическими кальмарами несколько увеличены. Например, у Lycoteuthis diadema диаметр глаза составляет более 15% длины тела. Увеличение глаз у глубоководных видов связано с уменьшением освещенности на глубинах.
Без преувеличения можно сказать, что глаза головоногих являются наиболее развитыми органами чувств, служащими для ориентации в пространстве. Только этим объясняется, по-видимому, отсутствие слепых головоногих моллюсков, среди обитающих даже на самых больших глубинах. Известен только один слепой вид головоногих моллюсков — глубоководный пелагический осьминог Cirrothauma murrayi. Если головоногие с поврежденными органами равновесия (статоцистами) сохраняют способность правильно ориентироваться при помощи глаз, то ослепленные всегда погибают от голода.
У некоторых глубоководных форм, в частности у представителей Cranchiidae (Galiteuthis, Bathothauma, Sandalops и др.), глаза сидят на стебельках различной длины.
У кальмаров семейства Histioteuthidae глаза асимметричны, левый в несколько раз больше правого. Установлено, что нормально развитый глаз приспособлен для зрения на глубине, а увеличенный — в приповерхностных слоях воды.
Строение глаз у кальмаров имеет важное систематическое значение. У подотряда Myopsida, к которому относятся неритические кальмары семейства Loliginidae. роговица сплошная и покрывает всю наружную поверхность глаза, лишь в переднем углу глаза сохраняется очень небольшое «слезное» отверстие. У подотряда Oegopsida, к которому принадлежат океанические кальмары, роговица имеет в центре широкое отверстие, размер которого почти равен диаметру глаза.
Различия в строении роговицы, по мнению ряда исследователей, имеют адаптивное значение и связаны с глубинами, на которых обитают кальмары. Вода, свободно проникающая через отверстие в роговице во внутренние полости глаза, способствует выравниванию давлений снаружи и внутри глаза и тем самым препятствует деформированию и разрыву оболочек глазного пузыря.
Представители Myopsida — прибрежные формы, тесно связанные с субстратом. Они, как правило, обитают вблизи берегов и не опускаются на сколько-нибудь значительные глубины. Обычно они встречаются на шельфе, на глубинах не более 200—250 м.
Кальмары подотряда Oegopsida встречаются в открытых районах моря на расстоянии до нескольких тысяч миль от берега. Вертикальное распространение Oegopsida не ограничивается поверхностными водами, они опускаются до глубин в несколько тысяч метров и совершают большие суточные вертикальные перемещения.
Недавно было установлено, что так называемые парольфакторные пузырьки — микроскопически малые органы, находящиеся в головном мозгу кальмаров между обонятельными долями и центральным мозгом (Loligo, Liocranchia, Bathothauma), — представляют собой активно функционирующие органы зрения — глаза внутри мозга! Возможно они регулируют фотопериодические реакции организма.
Руки и щупальца
Голова у кальмаров впереди заканчивается конечностями, радиально расположенными вокруг ротового отверстия и вытянутыми вдоль продольной оси тела. Обычно их пять пар — четыре пары рук и пара щупалец (ловчих рук). У некоторых кальмаров щупальца имеются только у личинок и молодых особей, а у взрослых они полностью редуцируются. Нумерация рук ведется со спинной пары, которая считается первой, верхне-боковая пара — второй, нижне-боковая — третьей и брюшная — четвертой.
Морфологически руки представляют собой сильно удлиненные конические выросты, более или менее треугольные в поперечном сечении. У многих кальмаров и прежде всего у быстроплавающих видов третья пара рук уплощена в спинно-брюшном направлении и образует выступающие по бокам рук горизонтальные кили, которые при плавании выполняют роль стабилизаторов и рулей. Наиболее сильно боковые кили развиты у нектонных кальмаров — Ommastrephidae, Onychoteuthidae и др.
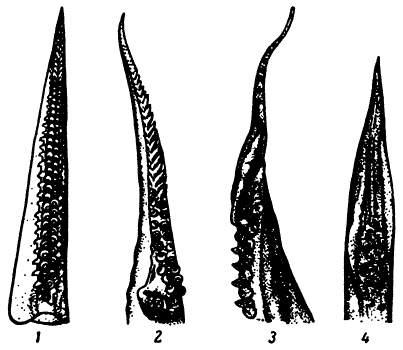
Гектокотилизированные руки самцов кальмаров: 1 — Sepioleuthis lessoniana; 2 — Loligo duvauceli; 3 — Pyroteuthis margarilifera; 4 — Symplectoicuthis oualaniensis (только конец руки)
Особенно хорошо развиты руки у быстроплавающих подвижных форм. У планктонных кальмаров руки обычно слабые, тонкие и короткие. Чаще всего самые мощные — вторая и третья пары рук, менее развиты первая и четвертая пары, но у Chiroteuthidae четвертая пара самая длинная.
Соотношение длины рук часто используется как систематический признак и обозначается формулой, в которой последовательность номеров пар соответствует соотношению длины рук. Например, формула рук 3.2.1.4 показывает, что наиболее длинные — руки третьей пары, а наиболее короткие — четвертой пары. Если какие-то пары рук имеют одинаковую длину, то в формуле между цифрами ставится знак равенства. Например, 2=3.4.1 показывает, что вторая и третья пары рук равны.
Внутренняя поверхность рук обычно по всей длине несет многочисленные присоски, а иногда крючки. С их помощью кальмары хватают и удерживают добычу. Присоски на руках располагаются, как правило, двумя продольными рядами и лишь иногда имеется четыре (Gonatidae) или иное число рядов. Крючки, заменяющие присоски у некоторых кальмаров (Enoploteuthidae, Octopodoteuthidae), представляют собой модификацию присосок.
Присоски кальмаров имеют твердое роговое (хитиновое) кольцо, наружный край которого гладкий, либо несет различной длины зубцы, характерные для тех или иных видов. У одних видов эти зубцы широкие и тупые (Loliginidae), у других узкоконические и острые (многие Oegopsida). Нередко центральный зуб рогового кольца присоски развивается значительно сильнее остальных. Дальнейшее развитие его может привести к превращению присоски в крючок (Gonatus).
Превращение присосок в крючья связывают с переходом кальмаров к питанию главным образом мягкотелой добычей — рыбами и кальмарами. Острые крючки, впивающиеся в мягкую ткань, обеспечивают более надежное удержание добычи. У осьминогов, питающихся в основном придонными ракообразными и двустворчатыми моллюсками, присоски не только никогда не превращаются в крючки, но даже не имеют твердых хитиновых колец.
Наиболее крупные присоски расположены на средней части руки, к основанию руки и к ее дистальному концу они уменьшаются в размерах.
Принцип действия присосок заключается в создании в момент присасывания пониженного давления внутри полости присоски. Присоска представляет собой чашу, ограниченную снизу с боков мускулистыми стенками. Объем полости присоски регулируется мускульными усилиями стенок. Механизм действия присосок можно сравнить с работой поршневого насоса: в качестве цилиндра выступает полость присоски, в роли поршня — дно ее. В нормальном состоянии все мышцы присоски (радиальные и кольцевые) находятся в расслабленном состоянии. Непосредственно перед присасыванием все мышцы сильно сокращаются, вследствие чего объем полости присоски уменьшается. В момент присасывания, как только край присоски плотно прижимается к поверхности схватываемого предмета, происходит моментальное расслабление всех мышц, за исключением лишь радиальных, вследствие чего объем внутренней полости увеличивается, а давление внутри нее падает. Разность давлений снаружи и внутри присоски обеспечивает ее плотное прикрепление.
Для отделения присоски от поверхности необходимо новое мышечное усилие, в результате которого выравнивается внешнее и внутреннее давление. Кальмары зачастую настолько прочно присасываются к жертве, что при резком движении последней разрывается стебелек присоски и присоска отрывается.
Измерения силы присосок у осьминогов показали, что присоска диаметром 2,5 мм способна удержать 57 г, а диаметром б мм — около 170 г.
Руки кальмаров обычно по всей длине оторочены тонкими прозрачными перепонками — защитными мембранами. Степень их развития различна у разных видов. В расправленном виде они поддерживаются поперечными мускульными перекладинами — выростами рук, а при плавании накрывают внутреннюю поверхность рук.
Между руками у кальмаров иногда (довольно редко) развивается тонкая перепонка, так называемая умбрелла, или «зонтик». Особенно хорошо развит зонтик у рода Histioteuthis. Кальмары этого рода, возможно, плавают с помощью зонтика подобно медузам.
У зрелых кальмаров часть одной руки, редко двух видоизменяется для выполнения половой функции. Чаще всего видоизменена брюшная левая или правая рука, иногда обе. Измененная часть руки называется гектокотиль, а процесс — гектокотилизацией.
Гектокотилизация у кальмаров выражается обычно в постепенном уменьшении размеров присосок от основания руки к ее дистальному концу и, наконец, полном их исчезновении. Стебельки, на которых сидят присоски, на гектокотилизированной части руки либо исчезают совсем, либо на дистальном конце руки превращаются в папиллы (усики). В отличие от некоторых осьминогов (Argonautidae) у кальмаров гектокотилизированная рука никогда не отрывается от тела.
Между третьей и четвертой парами рук почти у всех кальмаров имеется пара щупалец, которые иногда называют ловчими руками. У взрослых Octopodoteuthidae, Gonatopsis, Chaunoteuthis, Lepidoteuthis щупальца полностью редуцируются. Щупальца имеют тонкий стебель и расширенную дистальную часть. Это расширение называется булавой. Длина щупалец сильно варьирует даже у особей одного вида. Дело в том, что щупальца способны вытягиваться и сокращаться (стебель снабжен мощными продольными мышцами). Обычно при плавании стебель щупальца сокращается и у Loliginidae частично втягивается в специальную сумку (влагалище)» так что длина щупальца приближается к длине рук. Очень длинные щупальца у кальмаров семейств Architeuthidae и Chiroteuthidae, их длина иногда в несколько раз превышает длину тела.
На булаве щупалец располагаются присоски или крючки, а иногда и то и другое. Вооружение булавы всегда отлично от вооружения рук. Присоски на булаве располагаются в четыре, но нередко и в 8—12 продольных рядов, причем в двух средних рядах присоски крупные, а в краевых рядах мелкие. Самые крупные присоски находятся обычно в средней части булавы, к дистальному концу присоски сильно мельчают. В основании булавы, где она переходит в стебель, у многих кальмаров есть скопление мелких присосок и бугорков-кнопок, так называемый фиксирующий аппарат. Присоски и бугорки на левом и правом щупальцах расположены так, что бугорок одного щупальца соответствует присоске другого. При быстром плавании кальмары прижимают щупальца друг к другу и бугорки одного щупальца плотно входят в отверстия присосок другого. Оба щупальца прочно соединяются, причем булавы щупалец сохраняют подвижность и могут действовать наподобие клещей. Степень развития фиксирующего аппарата неодинакова. Например, у Chiroteuthidae фиксирующий аппарат отсутствует вообще, тогда как у Architeuthidae он занимает значительную часть стебля.
Рот, челюсти и радула
Между основаниями рук у головоногих моллюсков находится ротовое отверстие с мощными темно-коричневыми хитиновыми челюстями (мандибулами), напоминающими клюв попугаев. Благодаря такому сходству челюсти кальмаров также называют клювами. В отличие от клюва птиц, у головоногих нижняя челюсть заходит за верхнюю. Режущий край челюстей часто зазубрен. Строение клюва у кальмаров является важным систематическим признаком в определении видов.
Челюстями головоногие прокусывают твердые хитиновые панцири ракообразных, известковые раковины моллюсков, кости рыб, раскусывают добычу. Кальмары только раскусывают добычу, не перетирая ее; мелких животных они глотают целиком. Куски пищи транспортируются в глотку с помощью радулы, вооруженной семью, редко пятью рядами острых зубчиков, общее число которых достигает нескольких сотен. Она действует подобно конвейерной ленте.
Ротовое отверстие снаружи окружено тонкой кожистой пленкой, прирастающей к основаниям рук и к окружающей ротовое отверстие наружной губе. Это ротовая (буккальная) мембрана, или буквальная воронка. Она растянута на особых мускулистых тяжах (буккальных стрелах), расходящихся радиально от ротового конуса; их число может быть 6, 7 или 8, соответственно ротовая мембрана может быть шести-, семи- или восьмилучевая. Иногда ее лучи несут очень мелкие присоски, которым приписывают главным образом осязательные функции. Вообще чувствительные клетки многочисленны в присосках головоногих. У самок многих видов семейств Loliginidae, Ommastrephidae и других на ротовой мембране помещается одна или несколько специальных выстланных эпителием ямок — буккальных семеприемников. Это — резервуары, в которых сперма хранится до оплодотворения. Особенности устройства буккальной мембраны, в первую очередь число лучей, имеют систематическое значение.
Мантия
Туловище кальмаров со всех сторон окружено мантией, структура и форма которой сильно различаются у разных видов. Хотя обычно тело кальмаров плотное и мускулистое, встречаются формы с мягким студнеобразным телом. Столь значительные морфологические различия объясняются особенностями экологии этих видов, одни из которых ведут активный и подвижный образ жизни, тогда как другие малоподвижны.
Собственно форма тела кальмаров определяется формой мантии. Предпринимались попытки выделить наиболее характерные типы строения мантии головоногих моллюсков и классифицировать все многообразные формы тела Cephalopoda по девяти основным типам. Однако это деление несколько формально и не может считаться исчерпывающим.
Все быстроплавающие нектонные кальмары характеризуются сильно удлиненным изящным телом, что биологически вполне объяснимо. Только такая форма мантии позволяет этим кальмарам развивать высокие скорости при плавании, сводя до минимума величину сопротивления формы. Конец мантии нектонных кальмаров (Onychoteuthidae, Ommastrephidae) остро закруглен или приострен, что увеличивает обтекаемость тела.
У менее подвижных кальмаров форма тела претерпевает значительные изменения.
Наиболее характерные изменения мантии у планктонных видов — превращение ее в округло-мешковидную (Cranchia scabra), или, наоборот, чрезмерное удлинение, так что кальмар приобретает палочковидную форму (Doratopsis — личинка Chiroteuthis). Эти кальмары не имеют необходимости быстро перемещаться в толще воды, вследствие чего мантия у них потеряла обтекаемую форму.
У придонных Sepioteuthis мантия несколько уплощается, так что поперечное сечение тела становится эллипсовидным, причем брюшная поверхность более плоская, чем спинная. Форма мантии у придонных кальмаров оказывается промежуточной между формой мантии типичных пелагических кальмаров, с одной стороны, и каракатиц (Sepiidae), с другой. Биологический смысл уплощения тела у придонных видов нужно искать, по-видимому, в приобретении ими более совершенной маскирующей формы, помогающей надежно спасаться от врагов на дне. Переход к придонному образу жизни повлек за собой значительные целесообразные морфологические изменения.
Передняя часть мантии кальмаров имеет специальный замыкающий аппарат, наглухо закрывающий мантийную щель в момент выбрасывания воды из мантийной полости, так что вода может выходить только через воронку. Замыкающий аппарат различно устроен у представителей разных семейств. В типичном случае он состоит из трех пар хрящей-замков: одной дорсальной (затылочные хрящи) и двух боковых, располагающихся по сторонам основания воронки. При замыкании мантийной полости боковые мантийные хрящи плотно входят в соответствующие им хрящи-воронки. Затылочный хрящ мантии прочно соединен с затылочным хрящем головы.
Модификации замыкающего аппарата касаются прежде всего редукции вороночных хрящей и срастания вентрального края мантии с основанием воронки. Такое явление наблюдается у кальмаров семейства Cranchiidae.
У некоторых осьминогов мантия целиком срастается с головой, так что обмен воды осуществляется только через отверстие воронки. Эти осьминоги совсем утрачивают способность плавать при помощи маитийно-вороночного локомоторного аппарата.
Стенка мантии у кальмаров, как правило, толстая и мускулистая, особенно на брюшной стороне. Ее относительная толщина у нектонных Ommastrephidae достигает 4—5% длины мантии. Покровы мантии состоят из тонкого наружного эпителиального слоя (однослойный цилиндрический эпителий) и подстилающих его соединительнотканных слоев (собственно кожа). Поверхность кожи обычно гладкая. Кроме того, кожа кальмаров покрыта тонким слоем беловатой прозрачной слизи, который, выполняя роль гидродинамической смазки, еще более увеличивает гладкость.
В мускульном слое мантии наиболее развиты кольцевые мышцы. Кольцевой слой мышц занимает почти всю толщину стенки мантии. Сильное развитие кольцевых мышц связано с реактивным способом плавания кальмаров, при котором выталкивание воды из мантийной полости и создание пропульсивной силы достигается за счет сокращения именно этих мышц. Принцип действия кольцевых мышц мантии аналогичен работе сфинктеров позвоночных животных.
Значительно слабее кольцевых развиты радиальные и продольные мантийные мышцы. Радиальные мышцы пронизывают всю толщу стенки мантии, однако их объем не может идти ни в какое сравнение с объемом кольцевых мышц. Радиальные мышцы разделяют всю массу кольцевых мышц на отдельные поперечные кольца. Назначение радиальных мышц не совсем ясно; можно предполагать, что они принимают участие в управлении пограничным слоем при быстром плавании, создавая серии поперечных кольцевых волн при последовательных сокращениях мышц от хвоста к голове.
Продольные мышцы лежат тонким слоем на внешней поверхности кольцевых мышц. При их сокращении хвостовой конец мантии изгибается, что чрезвычайно важно для маневрирования кальмаров в толще воды.
Периодические сокращения мантийной стенки необходимы не только для создания реактивной тяги, они одновременно являются дыхательными движениями. В мантийной полости головоногих находятся жабры, нормальное функционирование которых невозможно без постоянного прокачивания свежих порций воды через мантийную полость.
Мантийная полость, таким образом, играет роль своеобразного резервуара или «камеры сгорания» гидрореактивного движителя, вследствие чего ее абсолютный объем определяет величину силы тяги, развиваемую животным. У быстроплавающих нектонных кальмаров объем мантийной полости составляет около половины объема тканей тела.
Плавники
У всех кальмаров мантия несет по бокам пару плавников, форма и размеры которых сильно различаются у разных видов. Обычно плавники прикреплены терминально — близ заднего конца мантии, но иногда они тянутся почти вдоль всего тела кальмаров (Sepioteuthis, Octopoteuthis, Thysanoteuthis). Напротив, у многих Crancbiidae плавники совсем маленькие. Размеры и форма плавников изменяются на протяжении жизни кальмара. У личинок и молоди плавники маленькие, короткие, округлые или лепестковидные, слабые. С возрастом они значительно удлиняются, становятся треугольными, ромбическими или сердцевидными, их мускулатура усиливается. Иногда возрастные изменения размеров и формы плавников столь значительны, будто плавники молодых и взрослых кальмаров принадлежат разным видам. Особенно заметны возрастные изменения плавников у быстроплавающих нектонных кальмаров, их личинки малоподвижны, и только с развитием мускулистых плавников кальмары получают возможность активного плавания.
Терминальное расположение плавников не случайно. Основная функция их — рулевая. Размещение плавников на конце тела, удаление их от центра тяжести дает возможность кальмарам пользоваться ими с наибольшим эффектом при минимальных затратах мускульной энергии. Значительную роль при поворотах наряду с плавниками играют гибкие руки, которые можно рассматривать как аналог передних рулей. Руки (передние рули) и плавники (задние рули) разнесены на противоположные концы тела, т. е. максимально удалены от центра тяжести, что усиливает эффект их совместного действия.
В результате одновременного действия плавников и рук возникает пара сил, вызывающая необходимый вращательный момент, который обеспечивает мгновенный поворот. Таким образом, кальмары имеют совершенную систему рулей как вертикальных, так и горизонтальных.
Воронка
Воронка представляет собой коническую трубку, расположенную на брюшной поверхности головы. Своим узким концом воронка направлена вперед, широким основанием — назад. Основание воронки срастается с туловищем и снизу закрыто мантией, передний же свободный конец ее лежит в особом углублении на брюшной стороне головы — вороночной ямке, которая может быть снабжена дополнительными складками или карманами.
Стенки воронки состоят из кольцевых мышц, которые могут регулировать диаметр отверстия воронки, а стало быть силу струи и скорость плавания кальмара. Чрезвычайная подвижность свободного конца воронки позволяет кальмарам легко и быстро менять направление движения. От заднего края воронки отходят хорошо развитые парные мускулы-депрессоры, которые при одновременном сокращении втягивают воронку. При сокращении одного из этих мускулов воронка разворачивается вниз и в сторону. Для прижимания воронки к голове служат мускулы-аддукторы.
Воронка функционирует как поворотное сопло гидрореактивного движителя. Разворачивая передний (свободный) конец ее, кальмар плывет в любом направлении. Маневренные качества кальмаров совершенны, без особых усилий кальмар меняет направление движения на обратное, двигаясь одинаково легко как головой, так и хвостом вперед.
Воронка кальмаров обычно снабжена клапаном. Вороночный клапан представляет собой языковидный вырост спинной стенки воронки и расположен возле самого выходного отверстия. Наиболее сильно развит он у нектонных кальмаров.
Сравнительно-экологический анализ кальмаров и лишенных клапана осьминогов дает основание рассматривать вороночный клапан как один из элементов гидрореактивного локомоторного аппарата. Эксперименты с ампутацией вороночного клапана у кальмаров показали, что его удаление лишает кальмаров способности к плаванию задним ходом — головой вперед, хотя изгибаемость воронки при этом остается прежней.
По-видимому функциональное значение вороночного клапана состоит в следующем. Из гидромеханики известно, что при движении жидкости в изогнутом канале на вогнутую стенку канала действует сила, величина которой определяется изменением количества движения массы жидкости при изменении направления ее движения. Другими словами, в изогнутом участке канала или трубы возникает своеобразный гидродинамический удар, который может вызвать повреждение стенки. Для предупреждения этого явления необходимо обеспечить дополнительную прочность изогнутых участков.
При плавании кальмаров головой вперед вороночный клапан расположен в изогнутой части воронки и можно предположить, что он является специальным образованием, обеспечивающим создание той необходимой механической прочности дорсальной стенки воронки, которая позволяет кальмарам развивать достаточно большие скорости при плавании задним ходом.
Увеличение механической прочности спинной стенки воронки за счет образования полужесткого клапана является более совершенным и прогрессивным признаком, нежели простое утолщение самой стенки, так как наряду с увеличением прочности сохраняется прежняя подвижность воронки, ее способность к изгибанию. Механическое объяснение функции вороночного клапана соответствует его развитию у кальмаров. У нектонных видов он хорошо развит, так как сила давления воды на стенку воронки у них, без сомнения, во много раз больше, чем у планктонных форм, например Cranchiidae, большинство которых не имеет клапана.
Источник: Г.В. Зуев, К.Н. Несис. Кальмары (биология и промысел). Изд-во «Пищевая промышленность». Москва. 1971