К настоящему времени получен целый ряд фактов, прямо или косвенно свидетельствующих о роли ДНК как первичной молекулярной структуры наследственности.
Перечислим основные аргументы в пользу того, что ДНК является основной макромолекулярной структурой наследственной информации, т. е. носителем программы функции гена и генотипа.
1. ДНК локализована в хромосомах или же в других способных к самовоспроизведению клеточных структурах.
2. Между количественным содержанием ДНК и числом хромосомных наборов в клетках найдена тесная постоянная связь. Так, в диплоидных ядрах содержится в два раза больше ДНК, чем в гаплоидных. Вместе с тем следует подчеркнуть, что указанное постоянство не является абсолютным. Содержание ДНК в ядре может несколько колебаться в зависимости от физиологической активности тканей.
3. Применение изотопной методики показало, что в неделящихся клетках ДНК практически не подвергается обновлению, т. е. не реплицируется, но в то же время активно контролирует синтез специфических белков.
4. Существует ряд специфических мутагенов на ДНК. ДНК более других химических компонентов клетки повреждается сильными мутагенами. Так, облучение микроорганизмов ультрафиолетом показало, что наиболее эффективные в отношении мутагенного действия лучи (имеющие длину волны около 260 ммк) наиболее интенсивно поглощаются ДНК. Высоким мутагенным действием на ДНК обладают также аналоги пуриновых и пиримидиновых оснований.
5. Твердо установленные факты передачи наследственной информации при трансформации, трансдукции и конъюгации посредством ДНК доказывают идентичность трансформирующего фактора и ДНК, а тем самым роль ДНК в наследственной передаче. Установлено, что почти чистый препарат нуклеиновой кислоты вируса передает наследственные признаки.
6. Обработка ДНК in vitro дезоксирибонуклеазой полностью инактивирует трансформирующий фактор.
Как видно будет из последующего изложения, структура ДНК бактериальной хромосомы, фага и клетки высшего организма наилучшим образом соответствуют исполнению функции наследственной информации, ее записи и хранению. Модель ДНК, предложенная Ф. Криком и Дж. Уотсоном в 1953 г., может способствовать пониманию некоторых основных генетических явлений на молекулярном уровне: существование самого гена, его действие, репликации, мутирования и рекомбинации. Так как основным способом изучения наследственности и, следовательно, структуры гена является изучение механизма мутаций, то, прежде всего, рассмотрим, как модель строения ДНК помогает решить этот вопрос.
Если генетическая специфичность определяется последовательностью пар нуклеотидов в молекуле ДНК, то возникновение мутации должно быть связано с заменой нуклеотидов. Можно предположить четыре типа таких изменений: 1) замена одного или большего числа нуклеотидов, 2) изменение последовательности пар нуклеотидов, 3) удвоение одной или большего числа пар нуклеотидов, выпадение одной или большего числа пар нуклеотидов.
Фриз, рассматривая молекулярный механизм мутаций, делит химические агенты на две группы. К одной группе он относит агенты, действующие на нуклеиновые кислоты в момент их репликации, репликации, к другой — действующие в состоянии покоя. Автор доказательно отмечает два возможных пути влияния химических факторов.
1. Подавление нормального образования предшественников нуклеиновых кислот: например, азасерин, подавляющий синтез пуринов, оказывается сильным мутагеном; также является сильным мутагеном уретан, подавляющий синтез пиримидинов. С этой точки зрения представляет интерес изучение спонтанного мутационного процесса у биохимических мутантов с нарушениями биосинтеза компонентов ДНК.
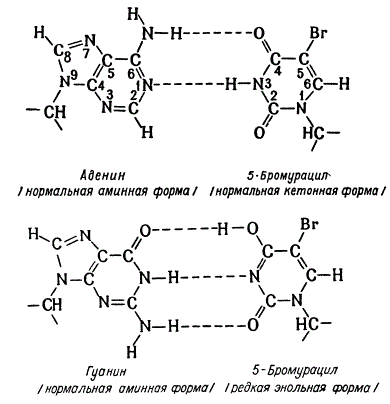
Схема спаривания аденина и гуанина с 5-бром-урацилом
2. Включение аналогов оснований. В ДНК и РНК нормально включаются свойственные им основания. В ДНК включаются А—Т, Ц—Г, а в РНК—А—У, Ц—Г. Установлено, что на место тимина в ДНК бактерий, фагов и клетки человека может быть включен 5-бромурацил и другие соединения урацила с галлоидами. 5-бромурацил в ДНК способен спариваться с аденином аналогично тимину. Иногда, но редко, происходит спаривание 5-бромурацила с гуанином. Замещение осуществляется при выращивании клеток на среде, в которой отсутствует тимин и куда добавляется 5-бромурацил. Во время репликации ДНК после расхождения комплементарных нитей аденин должен в норме спариваться с тимином, а цитозин с гуанином. Если при этом произойдет ошибка и вместо тимина к аденину пристроится 5-бромурацил, то в последующих репликациях ДНК 5-бромурацил может присоединить гуанин, который далее уже нормально спарится с цитозином. В результате пара оснований А—Т может уместиться на пару Г—Ц. В другом случае таким же образом пара Г—Ц может быть заменена парой А—Т.
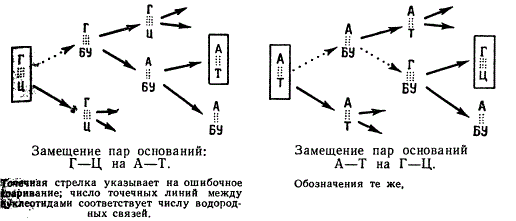
Замещение пар оснований
Сильный специфический мутаген азотистая кислота действует дезаминирования оснований. Дезаминированию в ДНК легче всего подвергается гуанин, затем цитозин и труднее всего аденин.
Результатом дезаминирования гуанина является ксантий, который образует водородные связи подобно гуанину, и потому на месте ксантина после репликации вновь оказывается гуанин, т. е. дезаминирование гуанина не выявляется. Некоторые авторы, правда, считают, что мутагенный эффект дезаминирования гуанина не выявляется в силу летальности этой комбинации. При дезаминировании цитозина образуется урацил, спаривающийся как тимин, и, образом, после репликации тимин оказывается на месте аденина. Дезаминирование аденина дает гипоксантин, спариваются как гуанин. Это приводит к появлению гуанина на месте.
Итак, замены оснований, вызываемые в ДНК азотистой кислотой, можно изобразить следующим образом:
Г → (К) → Г
Ц → (У) → Т
А → (ГК) → Г
В окончательном виде результат действия азотистой кислоты на выражается в замене пар: А—Т ⇔ Г—Ц.
Внешние агенты — мутагены могут изменять ход репликации ДНК и приводить к «ошибкам» последовательности нуклеотидов. Молекулярное исследование мутаций открывает возможности действия химическими и физическими агентами на выделенную in vitro. Измененная ДНК может быть введена путем трансформации в бактерии и выявлена затем как рекомбинация. Таким же образом можно использовать РНК вируса табачной мозаики (ВТМ). В обоих случаях удается точно учитывать реакции между применяемым химическим агентом и молекулами ДНК и РНК.
Изменение нуклеотидного состава ДНК и РНК приводит к мутациям генов, которые при включении ДНК в геном бактерии (в результате трансформации) и РНК в частицу ВТМ возможно учитывать количественно. При таких условиях опыта одно звено в цепи исследования мутагенеза, т. е. изменение ДНК и РНК, возможно проводить в пробирке, т. е. под контролем экспериментатора, а конечное проявление мутации учитывать по фенотипу живой клетки.
Таким образом, применение химических мутагенов, обладающих молекулярной специфичностью, т. е. способных вызывать определенные изменения в молекуле нуклеиновой кислоты, позволило доказать, что мутация есть нарушение, нуклеотидной последовательности в ДНК (реже РНК) гена.
Представление о нуклеиновых кислотах как носителях наследственной информации позволило выдвинуть ряд гипотез, объясняющих механизм рекомбинации сцепленных генов, т. е. механизм кроссинговера. Различные варианты этих гипотез сводятся к идее, сформулированной в 1963 г. X. Уайтхаузом. В основу предполагаемого механизма положено наиболее характерное свойство молекулы ДНК, а именно способность образовывать водородные связи между комплементарными основаниями А—Т и Г—Ц. Рассмотрим гипотезу в общей форме.
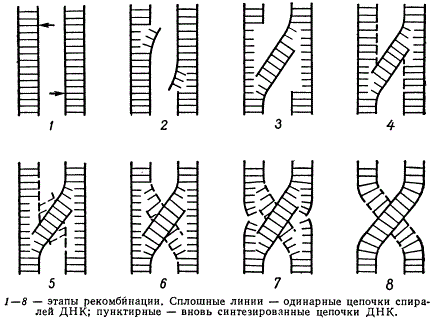
Возможный механизм рекомбинации на молекулярном уровне (стрелками указаны точки первоначальных разрывов)
Весь процесс обмена участками хроматид двух гомологичных хромосом, считая, что каждая хроматида представлена одной биспиральной молекулой ДНК, можно разбить на несколько этапов:
- разрыв одной цепи спирали ДНК в близких, но не гомологичных точках обеих молекул ДНК
- разрыв водородных связей между комплементарными цепочками ДНК вблизи разрывов и образование новой биспиральной «гибридной» структуры молекулы ДНК
- локальный синтез ДНК на месте отошедших цепочек
- расхождение вновь синтезированных цепочек в стороны и их комплементарное соединение
- разрыв цепочек, не претерпевших рекомбинацию и замыкание разорванных концов
Все этапы процесса происходят в ранней профазе мейоза, т. е. уже после того, как закончилась редупликация хромосом.
Предложенный механизм рекомбинации удовлетворительно объясняет большинство экспериментальных фактов, полученных на различных генетических объектах.