Первоисточником наследственной изменчивости является мутационный процесс.
В популяциях каждой породы и сорта диплоидных организмов спонтанно возникают разнообразные мутации. Под «защитой» доминантных генов дикого типа эти мутации накапливаются и сохраняются в гетерозиготном состоянии. По мере повышения концентрации мутантных аллелей в популяции возрастет вероятность их проявления в гомозиготном состоянии и увеличивается возможность комбинативной изменчивости. В природе судьбу этих мутаций определяет естественный отбор. При искусственном отборе сохранение таких мутаций определяет селекционер.
Породы и сорта отличаются по своим наследственным особенностям от диких предков. Если одомашнить дикие формы и создать им благоприятные условия произрастания или содержания, то они не воспроизведут продуктивности и качеств своих «культурных» сородичей. Как бы мы ни кормили банкивских кур, мы не смогли бы без длительного многолетнего отбора получить у них годовую яйценосность в 300—365 яиц, а также имеющееся у пород кур разнообразие по окраске пера, живому весу, скороспелости и т. д. Так же нельзя у муфлона или архара без длительной селекции получить шерсть типа мериносовой, у лошади Пржевальского — признаки современных скакунов с их разнообразными экстерьерными особенностями.
На основе искусственного отбора спонтанных мутаций и их комбинаций путем скрещиваний, при соответствующих условиях содержания и кормления человеку удалось в ряду многих сотен поколений вылепить новые формы животных и растений.
Поиски спонтанно возникших мутаций И. В. Мичурин называл «кладоискательством». Это было верно для того уровня селекции, когда не были изучены закономерности мутационного процесса и не были разработаны методы получения мутаций. Генетика выводит селекцию из состояния кладоискательства к сознательному овладению закономерностями мутационного процесса и дает в руки селекционера методы искусственного получения мутаций как у растений, так и у животных.
В качестве примера рассмотрим историю создания «сладкого» люпина. Все виды люпина имеют семена, содержащие ядовитые алкалоиды, а корни этих растений являются носителями клубеньковых азотфиксирующих бактерий. Поэтому долгое время люпин считался непригодным на корм скоту и использовался только на зеленое удобрение. У других видов бобовых существуют формы с безалкалоидными семенами, и, исходя из закона гомологических рядов, можно было полагать, что подобные мутации должны встречаться и у люпина. Для обнаружения мутантных растений сладкого Люпина была применена упрощенная методика определения безалкалоидных семян, что дало возможность оценить огромный материал и найти растения с безалкалоидными семенами. Однако найденные растения сладкого люпина имели другое отрицательное качество, а именно: створки боба у них раскрывались до уборки урожая, и семена осыпались. В дальнейшем среди 10 млн. проверенных растений все же было найдено одно растение с нераскрывающимися створками, но у этого растения при созревании опадали бобы. Размножив потомков этого растения, среди них стали искать мутантные формы с неопадающими бобами, и такие растения были найдены. Затем селекция сладкого люпина шла в направлении отбора форм с семенами, имеющими высокое содержание белка и прорастающими без периода покоя, а также растений с высоким положением соцветия, хорошей урожайностью и т. д. Все этапы этой работы строились исследователями на подборе соответствующих мутантов, встречаемость которых можно было предположить, согласно закону гомологии наследственной изменчивости.
Изучение мировых растительных ресурсов дало возможность Н. И. Вавилову выявить мировые очаги (центры происхождения) важнейших культурных растений. На основании определения ареала вида, изучения общей системы изменчивости с применением закона гомологических рядов и географической изменчивости сортового и видового разнообразия Н. И. Вавилов установил первичные и вторичные центры происхождения культурных растений. Первичные центры связаны с древними очагами цивилизации и местом первичного возделывания и селекции растений, а вторичные — с последующими периодами культуры земледелия. Такой подход к исследованию исходного материала Н. И. Вавилов назвал ботанико-географическим методом.
Было установлено, что наследственная изменчивость возделываемых растений определяется также историей их происхождения: популяции растений в центрах происхождения имеют наибольшую генетическую изменчивость.
Н. И. Вавилов выделил восемь мировых центров (очагов) происхождения культурных растений. Крупнейшим очагом земледелия и происхождения многих культурных растений, например хлебных злаков и других зерновых и бобовых (просо метельчатое, гаолян, гречиха, соя), является китайский; индийский очаг — родина риса, сахарного тростника, цитрусовых; среднеазиатский — мягкой пшеницы, гороха, бобов и других культур. Переднеазиатский очаг включает большое разнообразие видов пшениц и форм ржи; здесь находится мировой потенциал плодоводства. Средиземноморский, абиссинский, южно-мексиканский, южноамериканский очаги происхождения также имеют большое разнообразие специфических форм.
Знание очагов происхождения облегчает селекционерам поиски необходимых форм в местах их происхождения: именно здесь наиболее вероятны ценные генотипы.
В настоящее время особенно сильно возрастает роль искусственных мутаций в селекции. С открытием сильнодействующих мутагенов для селекции растений, микроорганизмов и насекомых открылись принципиально новые источники повышения изменчивости. На первых порах к практическому использованию индуцированных мутаций был проявлен скепсис даже среди виднейших генетиков и селекционеров. Этот скепсис объяснялся не только тем, что получаемые мутации часто носят патологический характер, но и отсутствием методов использования мутаций в селекции. Однако по мере изучения индуцированных мутаций положение изменилось, и применение ионизирующих излучений в селекции растений и микроорганизмов привело к созданию нового раздела в селекции — радиационной селекции. Хотя этот термин сам по себе неудачный, тем не менее, он получил распространение.
К числу пионеров применения ионизирующей радиации для целей селекции относятся наши советские генетики: Г. А. Надсон — в работах на микроорганизмах, Л. Н. Делоне и А. А. Сапегин — в селекции зерновых культур. Они применили ионизирующие излучения еще в 20-х и в начале 30-х годов. В те же годы сходные работы проводились в США Стадлером. Однако эти исследования были в то время оставлены без достаточного внимания.
Применяемые виды излучения могут быть различными: рентгеновы лучи, быстрые и медленные нейтроны и гамма-лучи. В настоящее время устраиваются специальные участки — «гамма-поля», где устанавливается на длительное время источник гамма-лучей с использованием Со60. Вокруг источника высаживают растения.
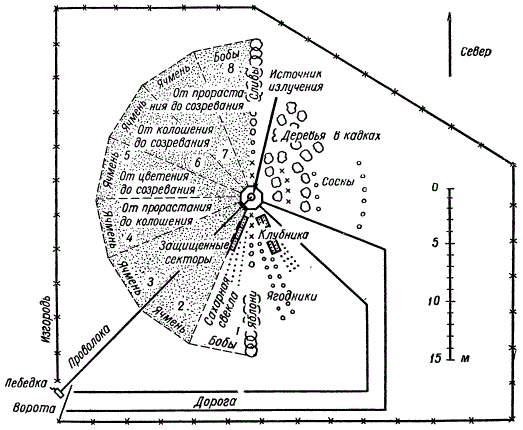
Примерный план участка гамма-поля
В радиационной генетике установлен ряд положений, имеющих важное значение для радиоселекции:
- покоящиеся и сухие семена выдерживают значительно более высокие дозы облучения, чем прорастающие семена и вегетирующие растения;
- покоящиеся растительные клетки менее чувствительны к ионизации, чем делящиеся и меристемные;
- хранение семян после облучения ведет к возрастанию числа хромосомных перестроек;
- разные виды и даже сорта растений имеют различную устойчивость к ионизации (горчица и другие крестоцветные, а также лен более устойчивы, чем бобовые, некоторые злаки и другие растения);
- полиплоидные растения более радиоустойчивы к ионизации, чем диплоидные;
- условия выращивания растений влияют на их радиочувствительность и выход мутаций.
У высших растений облучению подвергают семена, почки и пыльцу. Выбор стадии облучения зависит от биологии объекта и метода обнаружения мутаций. При возникновении рецессивной мутации в пыльцевом зерне, участвовавшем в оплодотворении, все клетки развивавшегося растения будут нести эту мутацию в гетерозиготном состоянии. У самоопылителя уже в следующем поколении можно выявить возникшую мутантную форму, а в последующих поколениях ее можно выделить и размножить. У перекрестноопыляющихся растений выявление рецессивных мутаций более трудоемко и требует не менее трех поколений.
Методы обнаружения мутантов после облучения семян иные, чем при облучении пыльцы, так как зародыш семени представляет многоклеточное образование, а мутации могут возникнуть лишь в отдельных соматических клетках. Поэтому выросшее из такого семени растение будет нести мутацию только в части тканей, берущих начало из измененной клетки. Такое растение окажется химерным; данное поколение принято обозначать как Х0. У пшеницы, ячменя и других злаков самоопылителей в пределах одного растения Х0 возникшая мутация может проявиться лишь в некоторых колосьях, а чаще лишь в одном или в нескольких зернах одного колоса.
Доминантная мутация может проявиться в первом же поколении, т. е. в Х1 а рецессивная окажется в Х1 в гетерозиготном состоянии и не проявится. Поэтому зерновки высевают отдельно по колосьям, используя по 15—20 семян из 2—5 нормально развитых колосьев. В течение вегетации растений Х2 среди них проводится выявление мутаций. Часть зерновок может оказаться носителями рецессивной мутации в гомозиготном состоянии, часть — в гетерозиготном. С мутантных растений Х2, представляющих селекционный интерес, собирают зерновки и вновь высевают для проверки и размножения в Х3, а затем для получения размноженной линии в Х4, Х5 и т. д. На рисунке приводится одна из возможных схем генетического анализа мутаций, возникающих после облучения на разных фазах развития растения.
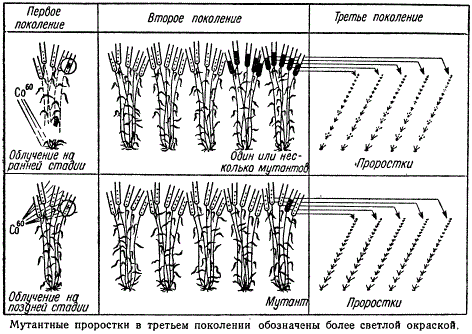
Схема генетического анализа мутаций, возникающих после облучения на разных фазах развитии растения
В селекции вегетативно размножающихся культур применяют облучение почек и побегов с целью получения у них доминантных соматических мутаций.
Успехи, достигнутые при использовании ионизирующих излучений в селекции, являются весьма перспективным началом. В последние годы в ряде стран (ГДР, Швеции, Канаде, США) уже получены промышленные сорта неполегающего ячменя, горчицы, рапса масличного. Созданы хозяйственно ценные формы яровой пшеницы, овса, ячменя, гороха, сои, арахиса, фасоли, люпина, клевера, томатов и др. Особенно важно отметить, что полученные в настоящее время мутанты служат важным источником изменчивости у зерновых культур для отбора по свойствам устойчивости к грибным заболеваниям: к мучнистой росе, стеблевой и листовой ржавчине. Получение хозяйственно ценных форм, устойчивых к различным заболеваниям, является одним из верных путей повышения урожайности растений.
Ярким доказательством важной роли индуцированных мутаций служат успехи в селекции продуцентов антибиотиков, аминокислот, витаминов. Как известно, продуцентами антибиотиков являются плесневые и лучистые грибы. Селекция по повышению активности продуцентов антибиотиков и витаминов группы В основывалась на отборе штаммов из природных популяций. Но отбор на основе изменчивости природных популяций вскоре исчерпал свои возможности, и перед экономикой ряда стран, в том числе и нашей, возникла альтернатива: либо значительно расширять фармацевтическую промышленность, либо находить методы повышения активности штаммов продуцентов антибиотиков. Так как второй путь более экономичен, то генетики и селекционеры направили свои усилия на получение индуцированных мутаций.

Продуктивность штаммов продуцентов антибиотиков и витаминов, выделенных из индуцированных мутантов
Приводятся сравнительные данные по активности исходных и новых штаммов продуцентов антибиотиков, полученных в лаборатории селекции ВНИИ антибиотиков под руководством С. И. Алиханяна. Мутации в этих опытах вызывались химическим веществом (этиленимином), рентгеновыми (Х-лучами) и ультрафиолетовыми лучами, а также комбинированным воздействием. Отбор мутаций позволил в короткий срок повысить продуктивность штаммов.
Успех использования индуцированных мутантов в селекции зависит от многих моментов: во-первых, от способа их получения, во-вторых, от разработанности методов обнаружения, объективности и доступности методики оценки мутантов, в особенности по биохимическим и физиологическим признакам, в-третьих, от изученности методов селекции мутантных форм, в-четвертых, от количества организмов, которые необходимо проанализировать для обнаружения нужных мутаций. Несмотря на многие трудности использования искусственных мутаций, это направление селекции раскрывает большие перспективы в деле создания новых форм растений, животных и микроорганизмов.
В селекции малоплодных животных получение индуцированных мутаций пока весьма ограничено. У ряда многоплодных позвоночных животных и у насекомых (шелкопряд, пчелы) получение мутаций может иметь практическое значение.