Для формирования нового организма у большинства растений требуется слияние двух половых клеток, или гамет — мужской и женской. Гаметы у покрытосеменных развиваются в специализированных органах цветка — тычинках и завязи.
Тычинки, или микроспорангиефоры, закладываются в конусе нарастания цветочного побега в виде бугорков. Вскоре после того как обозначился бугорок, в нижней его части начинает дифференцироваться тычиночная нить, в верхней — пыльник. Дифференциация тычинки на пыльник и тычиночную нить происходит через некоторое время после образования тычиночного бугорка. Если проследить развитие пыльника на поперечных срезах, можно убедиться, что округлые вначале очертания зачатка пыльника вскоре изменяются и становятся четырехугольными, затем интенсивное разрастание ткани зачатка с двух сторон делает его двулопастным. Между лопастями пыльника дифференцируется особая ткань, называемая связником, или коннективой. В результате дальнейшего неравномерного разрастания пыльник становится четырехлопастным.
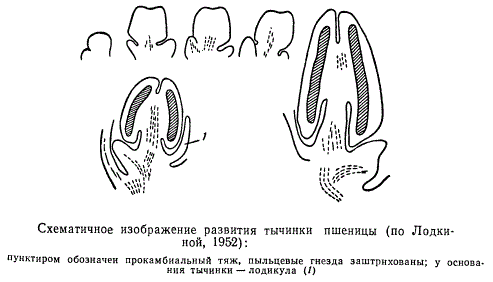
Расчленение пыльника на лопасти сопряжено с образованием в каждой лопасти по (пыльцевому гнезду — микроапорангию. Таким образом, типично сформированный пыльник состоит из четырех гнезд — микроспорангиев. Число гнезд у разных растений варьирует, некоторые гнезда могут зарастать или, наоборот, вместо четырех гнезд у некоторых растений образуется большее их число.
На ранних фазах развития пыльника пыльцевые гнезда состоят из меристематических клеток, окруженных снаружи эпидермальным слоем. Вскоре субэпидермальные клетки, которые можно уже рассматривать как клетки первичного археспория, делятся, отделяя кнаружи париетальные (пристенные) клетки, а ковнутри — клетки вторичного археспория. Клетки вторичного археспория могут размножаться митотически, увеличивая число себе подобных, или они непосредственно становятся спорогенными — материнскими клетками микроспор. Париетальные клетки также делятся, формируя тем самым стенку пыльника, которая у разных растений может состоять из разного числа слоев.
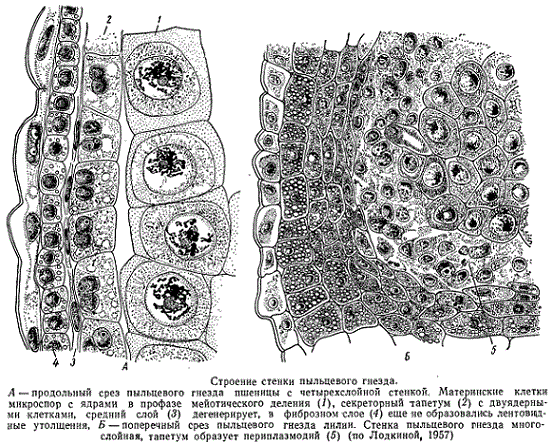
Внутренний слой стенки пыльника всегда представляет собой один или несколько рядов тапетальных, или выстилающих, клеток. У одних растений этот слой отчленяется при делении париетальных клеток, а у других — является производным вторичного археспория. Он непосредственно примыкает к материнским клеткам микроспор и состоит из таблитчатых, обычно дву- или многоядерных клеток. Различаются разные типы тапетального, или выстилающего, слоя: секреторный и амебоидный. При секреторном типе клетки тапетума сохраняют свою индивидуальность; при амебоидном, который иначе называют периплазмодием, оболочки клеток разрушаются, а протопласты сливаются в общую массу. Если исчезают оболочки всех клеток и все протопласты сливаются, то образуется так называемый типичный периплазмодий; если же исчезают не все оболочки и протопласты сливаются неполностью, то образуется нетипичный или неполный периплазмодий. Секретом тапетальных клеток служит желеобразная или слизистая масса, которая окружает микроспоры.
Секреторный тапетум изображен на рисунке, который показывает его состояние при развитии микроспор у пшеницы. Все клетки тапетума двуядерные. На рисунке изображен пример типичного периплазмодия из тапетума пыльника лилии.
Значение тапетума понятно — содержимое его разрушающихся клеток и выделяемый им секрет идет на питание развивающихся микроспор.
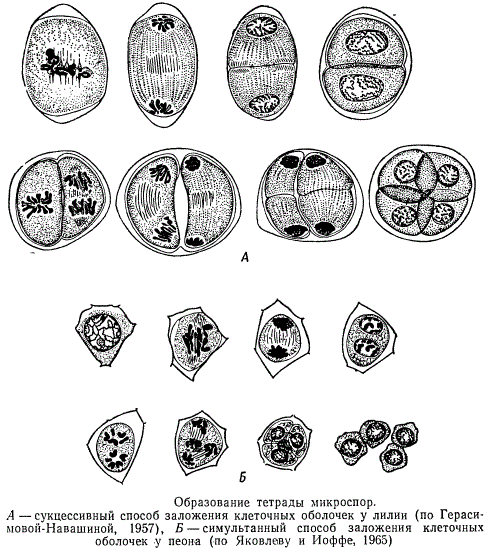
Достигнув некоторого предела в своем разрастании, материнские клетки микроспор начинают делиться, причем каждая клетка делится дважды, образуя 4 клетки, или тетраду, а каждая из вновь образовавшихся клеток тетрады дифференцируется в отдельную клетку, или микроспору. Деление материнских клеток микроспор отличается от обычного митотического деления тем, что число хромосом в дочерних клетках уменьшается вдвое, происходит редукционное, или мейотическое деление. Таким образом, из первоначально диплоидной, т. е. имеющей двойное число хромосом, материнской клетки образуются гаплоидные микроспоры с половинным числом хромосом. В результате этого процесса возникают качественно новые клетки, которые в дальнейшем дадут начало специфически мужским половым клеткам — спермиям.
Заложение клеточных оболочек в процессе редукционного деления может происходить двояко: у одних растений клеточные оболочки закладываются после каждого мейотического деления; такой способ называется последовательным, или сукцессивным, и встречается, например, у злаков, лилейных. У других растений клеточные оболочки, разделяющие будущие микроспоры, образуются лишь после второго мейотического деления; это так называемый одновременный, или симультанный, способ, встречающийся у лютиков, пеонов и многих других растений.
У представителей ряда семейств (рогозовых, вересковых, ластовневых и др.) микроспоры до опыления связаны в тетрады. В ряде случаев пыльцевые зерна соединены в группы по 8, 12, 16 и 32 микроспоры, как, например, в семействе мимозовых. У орхидных вся масса пыльцы каждой половины пыльника склеена особым веществом висцином в одну общую группу — поллиний.
После растворения оболочки тетрады микроспоры отделяются друг от друга; вокруг них формируются собственные оболочки, и прежде чем стать зрелыми пыльцевыми зернами, они проходят сложный путь развития. Каждая микроспора относительно сильно разрастается, в ней появляется большая вакуоль, оттесняющая клеточное ядро к периферии клетки. Ядро микроспоры и его ядрышко значительно увеличиваются, после чего микроспора приступает к делению, в результате которого образуются две клетки. В связи с периферическим положением ядра возникающая в процессе деления перегородка разделяет микроспору на две клетки неравной величины с ядрами и ядрышками также неравных размеров.
Меньшая линзообразная клетка, имеющая небольшое овальное, богатое хроматином ядро с маленьким ядрышком или без него, называется генеративной. Большая клетка, содержащая бедное хроматином ядро, называется вегетативной. Таким образом, первое деление микроспор является началом формирования мужского гаметофита. Цитоплазма генеративной клетки отличается от цитоплазмы вегетативной клетки густым содержимым. По количеству цитоплазмы в генеративной клетке разных растений различают «многоплазменные» и «бедноплазменные» генеративные клетки. Оболочка генеративной клетки очень тонкая, прозрачная, обладает высоким показателем преломления и не дает реакции на целлюлозу; такая оболочка легкопроницаема и облегчает обмен веществ между нею и вегетативной клеткой. Включения генеративной клетки составляют пластиды и хондриосомы. Вегетативная клетка, кроме того, содержит жир, крахмал, протеин и т. д.
Генеративная клетка одной своей стороной первоначально прилегает к оболочке микроспоры, но постепенно она отделяется от этой оболочки и передвигается внутрь микроспоры, где и располагается в цитоплазме вегетативной клетки, часто принимая веретенообразную или серповидную форму.
В процессе развития генеративная клетка тесно взаимодействует с вегетативной: расположение ее в глубине вегетативной наилучшим образом обеспечивает ей питание, так как она со всех сторон окружена цитоплазмой вегетативной клетки. В то время как вегетативная клетка растет и развивается за счет притока питательных веществ извне, из среды, в которую погружены микроспоры, генеративная клетка дифференцируется за счет питательных веществ вегетативной клетки.
В процессе дальнейшего формирования пыльцевого зерна ядро вегетативной клетки больше не делится, ядро же генеративной клетки и сама клетка делятся, образуя два спермия. У одних растений спермин (мужские гаметы) образуются в самом пыльцевам зерне (злаки, осоки, сусаковые, сложноцветные и др.); у других — при его прорастании — в пыльцевой трубке (лютиковые, норичниковые, лилейные и др.). В первом случае пыльцевое зерно трехклеточное, во втором — двухклеточное.
Долгое время дискутировался вопрос о том, что представляют собой спермии, — клетки или ядра. Сейчас уже достаточно данных в пользу клеточной природы спермиев, в которых содержится не только ядро, но и цитоплазма (правда, в весьма ограниченном количестве). Изображение спермиев в виде ядер, часто встречающееся в оригинальных работах, связано с тем, что цитоплазма их не всегда выявляется при применяемых обычно ядерных красителях, но может быть обнаружена при специальных методах окрашивания.
Ко времени раскрывания пыльника и рассеивания пыльцы пыльцевые зерна вполне сформированы и имеют две оболочки: внутреннюю — интину и наружную — экзину. У некоторых водных растений зрелые микроспоры имеют лишь одну оболочку.
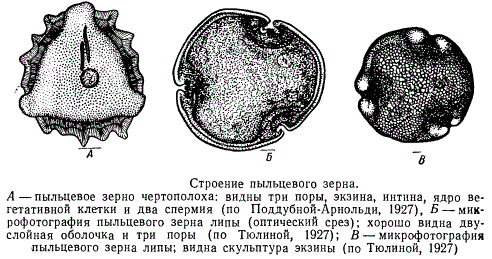
Интина представляет собой тонкую однородную пленку, состоящую преимущественно из пектиновых веществ. Экзина — довольно толстая, грубая, нередко слоистая, кутинизированная оболочка. Поверхность ее, как правило, обладает сложной скульптурой, весьма характерной для каждой систематической группы растений.
Постоянство этого признака широко используется в систематике для диагностических целей.
Экзина непроницаема или малопроницаема для находящихся вне ее веществ и весьма устойчива к различным воздействиям. Интенсивность обмена веществ зрелого, но еще не начавшего
прорастать пыльцевого зерна незначительна, так как после созревания оно впадает в более или менее продолжительное состояние относительного покоя. Для зрелого пыльцевого зерна характерны борозды и поры, т. е. неутолщенные места, а иногда сквозные отверстия, достигающие интины. Последние данные, полученные при помощи электронного микроскопа, говорят о том, что оболочка пыльцы, например некоторых злаков, пронизана каналами, через которые ее содержимое сообщается с окружающей средой.
Пыльца различных видов покрытосеменных растений различается не только структурой экзины, числом и характером отверстий для выхода пыльцевых трубок, но также своей величиной, формой и включениями. У одних растений, например у незабудки, пыльца очень мелкая, у других (тыква, мальва) — относительно крупная. Разумеется, между крайними пределами пыльцевых зерен существуют и промежуточные. Иногда размеры пыльцы значительно варьируют даже у разных форм одного и того же вида растения. Например, полиплоидные формы имеют значительно более крупные пыльцевые зерна и нередко большее число пор в них, чем соответствующие диплоидные и гаплоидные формы того же вида.
По своей форме пыльцевые зерна бывают округлыми, эллипсоидными, тетраэдрическими, бисквитообразными, удлиненными и т. д.
Основными запасными веществами пыльцы являются крахмал и сахар, реже жир и белок. Из белковых веществ пыльца содержит альбумин, глютамин и ряд аминокислот. В пыльце обнаружены каротиноиды. В пыльце и пыльцевых трубках обнаружен ряд ферментов: инвертаза, диастаза, каталаза, редуктаза, пектиназа, пепсин, трипсин, эрепсин, липаза и цитаза. Кроме того, пыльца содержит значительное количество физиологически активных веществ типа ростовых гормонов, стимулирующих прорастание и рост пыльцевых трубок.
Количество пыльцевых зерен в пыльниках различных видов растений весьма различно: у одних растений оно исчисляется десятками, у других — сотнями, у третьих — тысячами и сотнями тысяч. Обычно в цветке образуется значительно большее число пыльцевых зерен, чем, казалось бы, нужно для того, чтобы обеспечить оплодотворение семяпочек. Целесообразность образования большого количества пыльцы заключается в том, что благодаря этому обеспечивается оплодотворение при всех возможных, даже неблагоприятных условиях жизни организма, и создается большой запас генетически разнообразных мужских гамет, благоприятствующий избирательности при оплодотворении.
Высыпание созревшей пыльцы и ее рассеивание происходит благодаря вскрыванию пыльника, которое осуществляется разными способами. Большую роль в этом процессе играет так называемый фиброзный слой, или эндотеций, представляющий собой видоизмененный в процессе развития микроспорангия субэпидермальный слой его.
В клетках стенки молодого пыльника, расположенных непосредственно под эпидермисом, протопласт начинает сгущаться, в нем появляются зеленые пластиды (хлоропласты), в которых отлагаются зерна крахмала. В дальнейшем цитоплазма выделяет целлюлозу и гемицеллюлозу, откладываемые в каждой клетке в виде неравномерных утолщений ее стенки; хлоропласты при этом разрушаются. Характер клеток фиброзного слоя очевидно связан с его функцией, обычно вскрывание пыльника происходит в результате подсыхания и сокращения оболочек клеток фиброзного слоя. У некоторых семейств покрытосеменных растений типичный фиброзный слой отсутствует, у других имеются два и более слоев.
Обычно там, где должна образоваться щель пыльника, при его раскрывании формируется слой тонкостенных клеток, легко разрывающийся. Способы раскрывания пыльника многообразны: продольной или поперечной щелью, порами, клапанами и т. д. Наиболее распространено раскрывание продольной щелью.
При попадании зрелой пыльцы на рыльце пестика она начинает прорастать; содержимое пыльцевого зерна, одетое интиной, образует вырост, называемый пыльцевой трубкой. Прорастание пыльцевых трубок происходит через проростковые борозды, или поры (апертуры). Число борозд и пор у разных растений может быть неодинаковым и является, как и скульптура экзины, диагностическим признаком. Иногда поры снабжены «крышечками», открывающимися под давлением прорастающей пыльцевой трубки.
Несмотря на существование нескольких пор, содержимое пыльцевого зерна прорастает чаще всего лишь одной пыльцевой трубкой. При помощи пыльцевой трубки мужские половые клетки-спермии доставляются в зародышевый мешок, где они сливаются с элементами женского геметофита в процессе оплодотворения.
Пыльца может прорастать не только на рыльце, но и на искусственной питательной среде. Проращивание на искусственной среде дает возможность быстро определить степень ее фертильности, а также облегчает исследование пыльцы.
Скорость прорастания пыльцы и роста пыльцевых трубок различны у разных покрытосеменных растений, колеблясь от нескольких минут до многих часов, дней, недель и даже месяцев. Внешние условия, особенно температура и влажность, играют важную роль как при образовании, так и при прорастании пыльцы. Обычно высокая температура и низкая влажность ускоряют рост пыльцевых трубок, низкая температура и высокая влажность, наоборот, замедляют его. При этом огромное значение имеет и «возраст пыльцы»: чем она старше, тем медленнее прорастает и тем медленнее растут пыльцевые трубки.
Мужские гаметы большинства животных и растительных организмов способны к активному движению. Поэтому исследователей издавна интересовал вопрос, обладают ли также спермин покрытосеменных этой способностью или они (передвигаются пассивно, увлекаемые током цитоплазмы вегетативной клетки. Однако до настоящего времени не сложилось единого мнения по этому вопросу.
Что касается жизнеспособности пыльцы, то у разных растений она также неодинакова. У одних видов пыльца быстро теряет свою жизнеспособность и перестает прорастать; у других сохраняет фертильность более или менее продолжительное время. У большинства злаков пыльца жизнеспособна 1—3 суток, у кок-сагыза и пеона — 5, мака — 14, цикламена — 18, душистого горошка — 23, рододендрона — 47, нарцисса — 72, колхикума — 92, подсолнечника — 386 суток. Разумеется, сохранение пыльцой жизнеспособности в течение более или менее продолжительного времени связано с условиями хранения. Наиболее благоприятными условиями для хранения являются пониженная температура и умеренная влажность.
Женским половым органом, в котором развиваются женские гаметы — яйцеклетки, является пестик. В нем различают три части: завязь, столбик и рыльце. У подавляющего большинства растений пестик представляет собой сложное образование, сформированное благодаря срастанию нескольких составляющих его плодолистиков. Плодолистик соответствует тычинке и является мегаспорангиефором. В начале своего развития зачатки тычинок и плодолистиков очень сходны. Лишь спустя некоторое время начинается процесс их дифференциации; бугорки, нижерасположенные на конусе нарастания, приобретают специфические черты строения микроспорангиефора (этот процесс можно назвать маскулинизацией), вышерасположенные бугорки становятся мегаспорангиефорами (феминизация).
В образовании пестика может принимать участие разное число плодолистиков и они могут по разному между собой срастаться. В соответствии с этим основная нижняя часть пестика — завязь — может иметь различное число полостей, или гнезд. В полостях завязи образуются и развиваются семяпочки, или мегаспорангии, соответствующие пыльцевым гнездам, или микроспорангиям. В этих мегаспорангиях в процессе их дифференциации и формируются женские половые гаметы — яйцеклетки.
Место заложения семяпочек в завязи называется плацентой. Характер плацентации и ее эволюция определяются направлением эволюции самого гинецея. Имеются различные типы плацентации, но в основном они сводятся либо к ламинальному типу, когда семяпочки расположены на внутренней поверхности плодолистика, либо к сутуральному типу, когда семяпочки сидят вдоль швов плодолистиков.
Вскоре после того, как возникает бугорок семяпочки, начинается наружная и внутренняя его дифференциация: параллельно с ростом центральной части семяпочки, называемой ядром, или нуцеллусом, развиваются покровы ее, или интегументы. Чаще всего образуются два покрова — наружный и внутренний. Однако у многих растений семяпочки имеют только один покров и лишь у некоторых растений совершенно нет покровов — семяпочка голая.
 Помимо интегументов, у некоторых видов растений на семяпочках обычно сразу после оплодотворения (иногда до оплодотворения) образуются особые выросты, известные под различными названиями: ариллусы, кровельки, присемянники, карункулы (или микропилярные ариллусы), строфиоли и т. д. При созревании семян они принимают яркую окраску, но иногда остаются бесцветными.
Помимо интегументов, у некоторых видов растений на семяпочках обычно сразу после оплодотворения (иногда до оплодотворения) образуются особые выросты, известные под различными названиями: ариллусы, кровельки, присемянники, карункулы (или микропилярные ариллусы), строфиоли и т. д. При созревании семян они принимают яркую окраску, но иногда остаются бесцветными.
Все разнообразие форм этих образований можно свести к двум основным группам: ариллусы — образования, отходящие от семяножки (фуникулуса) в области рубчика, облегающие семя целиком или частично, не срастающиеся с семенной кожурой, и ариллоиды — различного рода разрастания наружного интегумента.
Термин ариллус часто употребляется в широком смысле, включая и ариллусы, и ариллоиды.
На верхушке семяпочки покровы, как правило, не срастаются друг с другом, образуя подобие канала, называемого пыльцевходом, или микропиле, служащим для прохождения пыльцевой трубки к зародышевому мешку.
Микропиле может быть образовано либо внутренним покровом (эндостом), либо наружным (экзостом), либо обоими вместе. В последнем случае часть микропиле, образуемая наружным покровом, может не вполне совпадать с частью, образуемой внутренним покровом, вследствие чего микропиле становится не прямым, а зигзагообразным. Не у всех растений микропиле представляет собой полый канал, у некоторых оно заполнено более или менее вытянутыми клетками и даже волосками. В некоторых случаях, когда покровы семяпочки полностью срастаются или когда нуцеллус выдается из покровов, микропиле отсутствует.
У ряда растений клетки, выполняющие канал микропиле, становятся железистыми или образуют слизистую массу, что, по-видимому, служит для питания пыльцевой трубки и облегчает прохождение пыльцевых трубок к зародышевому мешку.
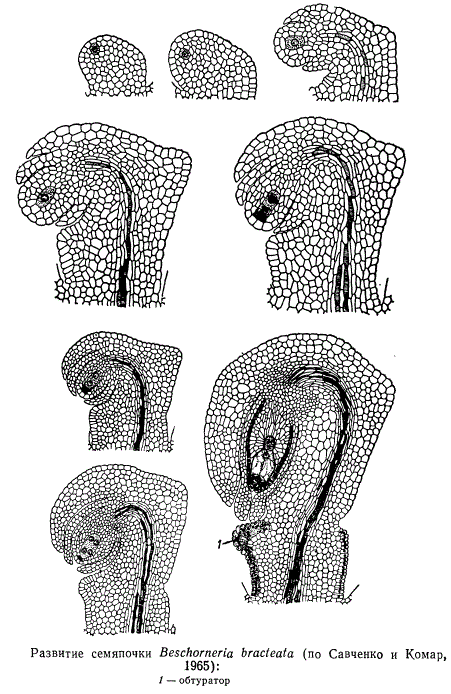
Иногда клетки наружных частей плаценты, внутреннего или наружного интегументов, нуцеллуса, фуникулуса или плодолистиков сильно разрастаются и вытягиваются в направлении микропиле, вследствие чего образуется так называемый обтуратор. Полагают, что обтуратор облегчает пыльцевым трубкам проникновение в завязь, являясь как бы мостом для продвижения пыльцевых трубок из столбика к микропиле семяпочки. Клетки обтуратора структурно напоминают клетки проводниковой ткани или железистых волосков.
Семяпочки соединены с плацентой при помощи более или менее длинной ножки, называемой семяножкой, или фуникулусом. Место перехода семяпочки в семяножку называется халазой, а вся нижняя часть семяпочки — соответственно халазальной. Верхняя часть семяпочки, примыкающая к микропиле, называется микропилярной частью.
По мере развития семяпочки клетки тканей нижнего и верхнего концов ее иногда претерпевают утолщения, одревеснение или опробкование; такие образования в нижней части семяпочки называются гипостазой, в верхней — эпистазой.
В результате резорбирующего действия зародышевого мешка гипостаза часто вдается в зародышевой мешок, образуя особую ткань, называемую постаментом. Особенно хорошо она выражена у представителей семейств лютиковых и липовых.
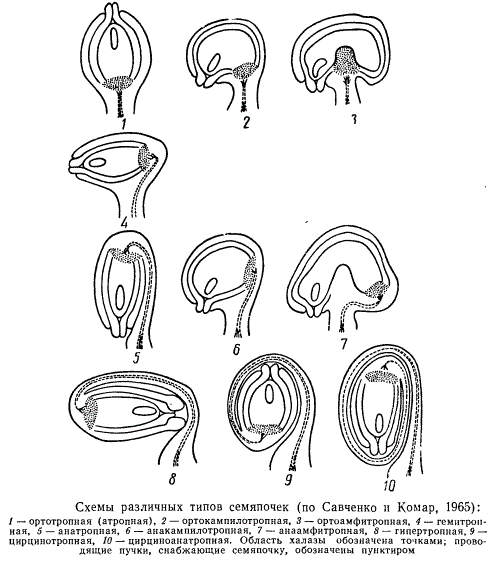
Форма, величина и количество семяпочек у разных групп покрытосеменных растений различны. Неодинаковы также характер и степень изгиба семяпочки в процессе ее развития по отношению к фуникулусу. В связи с этим различают разные типы семяпочек. Некоторые исследователи выделяют пять основных типов, другие приводят большее число, которое также, вероятно, не исчерпывает всего их разнообразия. На рисунке представлено схематичное изображение десяти типов семяпочек, характеристика которых кратко сводится к следующему:
- Прямые — ортотропные, или атропные, семяпочки. Микропиле находится на одной оси с семяножкой и на противоположном конце по отношению к халазе.
- Обращенные — анатропные семяпочки. Тело семяпочки повернуто на 180°, вследствие чего халаза и микропиле находятся на противоположных концах, а микропиле и рубчик расположены рядом.
- Полуповернутые — гемитропные, или гемианатропные. Семяпочка повернута по отношению к семяножке на 90°, микропиле и халаза находится на противоположных концах.
- Согнутые — кампилотропные семяпочки. Вследствие одностороннего разрастания нуцеллуса и интегументов семяпочка сильно изогнута и микропиле располагается почти на одном уровне с халазой. Среди них различаются орто кампилотропные семяпочки, у которых халаза и проводящий пучок, снабжающий семяпочку, располагаются на одной прямой оси с семяножкой, и анакампилотропные, у которых микропиле и халаза находятся на разных уровнях, а проводящий пучок поднимается по семяножке к халазе.
- Двусторонне согнутые — амфитропные семяпочки. Нуцеллус семяпочки перегибается по середине таким образом, что в продольном разрезе он приобретает подковообразную форму. Микропиле и халаза расположены на одном уровне или почти на одном уровне. Среди них различаются ортоамфитропные семяпочки, у которых халаза и проводящий пучок, снабжающий семяпочку, находятся на прямой оси с семяножкой, и анаамфитропные семяпочки, у которых проводящий пучок искривляется вслед за изгибом семяпочки и подходит к халазе.
- Сверхобращенные — гипертропные семяпочки. У таких семяпочек семяножка отходит от плаценты под прямым углом, затем сильно удлиняется и изгибается таким образом, что нуцеллус с интегументами поворачивается на 270°.
- Закрученные — цирцинотропные семяпочки. Сначала микропиле расположено на одной линии с основной осью семяпочки, затем семяпочка изгибается, полностью переворачивается и ее микропиле подобно прямой семяпочке снова оказывается направленным вверх, а ось нуцеллуса располагается параллельно семяножке.
- Закрученно-обращенные — цирциноанатропные семяпочки. Характеризуются тем, что семяножка настолько удлиняется, что ось нуцеллуса и интегументы делают полтора оборота. Таким образом, тело семяпочки поворачивается на 540°.
Семяпочки покрытосеменных растений значительно варьируют и по характеру нуцеллуса. У одних нуцеллус состоит из немногих клеток, и материнская клетка мегаспор обычно лежит непосредственно под эпидермисом — это так называемые тенуинуцеллятные семяпочки, примером которых могут служить семяпочки орхидных, первоцветных, сложноцветных и пр. У других нуцеллус состоит из многих клеток, а материнская клетка мегаспор отделена от эпидермиса одним или несколькими слоями клеток — это так называемые крассинуцеллятные семяпочки, характерные для розоцветных, некоторых лютиковых, ивовых и др.
Семяпочки покрытосеменных растений могут быть очень мелкими, различимыми лишь при помощи микроскопа, или более крупными, обнаруживаемыми невооруженным глазом. Обычно чем более мелки семяпочки, тем их больше в завязи. Количество же семяпочек в завязях различных представителей покрытосеменных колеблется от одной до многих сотен и даже тысяч. У ряда видов орхидей в одной завязи обнаружено до 2 млн. семяпочек.
По мере дифференциации семяпочки на поверхности нуцеллуса и покровов образуется эпидермис из клеток с более или менее утолщенными оболочками, которые в процессе образования семенной кожуры могут ослизняться, одревесневать, опробковевать, кутинизироваться и т. д. Эпидермис покровов наиболее устойчив и в построении семенной кожуры играет наиболее существенную роль. Паренхима нуцеллуса и покровов по мере развития семени обычно облитерируется. Тонкие клеточные оболочки их, состоящие из целлюлозы и пектиновых веществ, не препятствуют прохождению через них питательных растворов, направляющихся к центру семени.
В паренхиме нуцеллуса и покровов, выполняющей преимущественно питательную функцию, нередко можно обнаружить пластиды, крахмал, сахар, жир, белок, аминокислоты, кристаллы минеральных солей, дубильные вещества, а также пигменты и среди них — хлорофилл.
Элементы проводящей системы семяпочки — сосудистые пучки— тянутся от семяносца по семяножке вплоть до халазы. У многих покрытосеменных сосудистые пучки в халазе и оканчиваются, у некоторых же видов растений они проникают также в интегументы и очень редко в нуцеллус. Семяпочки со слаборазвитым нуцеллусом, который быстро разрушается, обычно имеют так называемый эндотелий, или интегументальный тапетум — слой клеток таблитчатой радиальноудлиненной формы, наполненных густым содержимым с сильно хроматизованными ядрами.
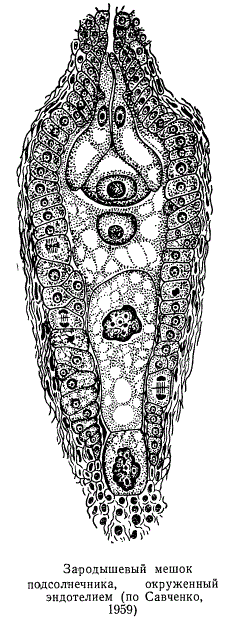
Эндотелий образуется из внутреннего эпидермиса покрова и примыкает непосредственно к зародышевому мешку, окружая его. По-видимому, он выполняет функции, связанные с питанием зародышевого мешка, т. е. является посредником при передвижении питательных веществ из покрова в зародышевой мешок. Возможно, на ранних стадиях развития семяпочки эндотелий является питающим слоем, а на более поздних, по мере созревания зародыша — защитным слоем. Кроме того, эндотелию свойственны функции и железистого образования.
По мере развития семяпочки в нуцеллусе, обычно непосредственно под его наружным слоем: дифференцируется клетка, выделяющаяся среди прочих своей величиной, размерами ядра и ядрышка, а также густой цитоплазмой. Эта клетка представляет собой клетку первичного археспория; в дальнейшем она либо непосредственно превращается в материнскую клетку мегаспор, либо делится и образует париетальную и спорогенную клетки. Нередко образуется не одна, а две и даже несколько клеток археспория, вследствие чего и женский археспорий может быть дву- и многоклеточным.
Вскоре после своего формирования материнская клетка мегаспор, имеющая диплоидное число хромосом, претерпевает два последовательных мейотических деления, в результате которых аналогично образованию тетрады микроспор образуется тетрада гаплоидных мегаспор. Возникают качественно новые клетки, которые дадут начало женской гамете — яйцеклетке. Перегородки между мегаспорами образуются последовательно. В зависимости от направления перегородок расположение клеток в тетраде различно: наиболее часто встречается линейное и Т-образное расположение мегаспор. При линейном расположении мегаспоры располагаются друг за другом, при Т-образном — две клетки расположены горизонтально, а две — перпендикулярно к ним.
У громадного большинства покрытосеменных растений образуются четыре клетки мегаспор, из которых в преобладающем числе случаев нижняя увеличивается в размерах, превращаясь в зародышевый мешок, а три верхние отмирают. Их остатки в виде бесформенных сгустков можно еще долго видеть на разных стадиях развития зародышевого мешка. Последний, развиваясь и увеличиваясь в размерах, постепенно их оттесняет и ассимилируем.
Описанный выше мегаспорогенез, при котором дальнейшее развитие претерпевает лишь одна мегаспора, приводит к образованию так называемого моноспорического зародышевого мешка. У некоторых растений в формировании женского гаметофита — зародышевого мешка — принимает участие большее число мегаспор — 2 или 4. В таких случаях образуются соответственно биспорические или тетраспорические зародышевые мешки.
Внутри каждого из этих трех типов женского гаметофита имеется значительное число вариаций, обусловленных разным числом делений ядер, приводящих к формированию зрелого зародышевого мешка. В результате зародышевый мешок, или женский гаметофит, у разных покрытосеменных растений может состоять из неодинакового числа ядер, в дальнейшем же в соответствии с числом этих ядер формируется различное число клеток.
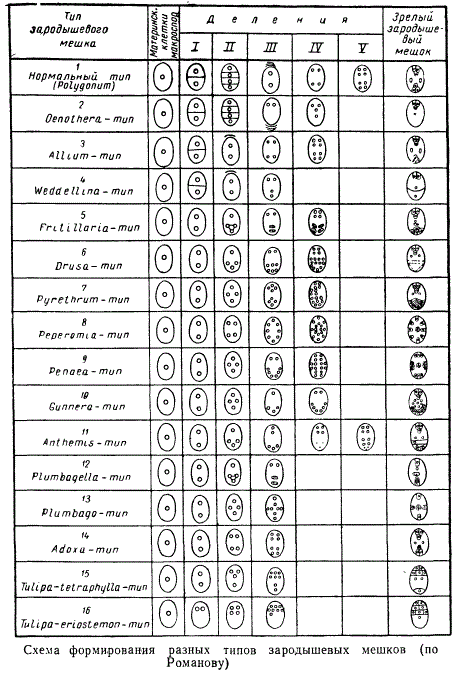
Не разбирая все имеющиеся вариации зародышевых мешков, рассмотрим лишь наиболее широкораспространенный случай — так называемый нормальный тип зародышевого мешка, обозначаемый как Polygonum-тип — по названию растения, у которого он впервые был открыт.
Как уже отмечалось, из четырех мегаспор, образовавшихся в результате редукционного деления, три разрушаются и лишь одна (обычно это халазальная мегаспора) увеличивается в размере, превращаясь в одноядерный зародышевый мешок.
Вскоре единственное ядро формирующегося зародышевого мешка делится и он становится двуядерным. Оба ядра расходятся к противоположным полюсам, а в центральной части разросшейся клетки возникает разделяющая их вакуоль. Затем ядра снова делятся и двуядерная стадия зародышевого мешка сменяется четырехъядерной. В свою очередь каждое из четырех ядер снова делится — возникает восьмиядерная стадия. В результате этого последнего деления у каждого полюса формирующегося зародышевого мешка возникает по группе из четырех ядер, разделенных обширной вакуолью. В период вышеописанных делений зародышевый мешок разрастается, принимая различную у разных видов форму: округлую, овальную, изогнутую.
Наряду с увеличением размеров зародышевого мешка идет разрастание семяпочки в целом. Рост и развитие зародышевого мешка осуществляются преимущественно за счет окружающих его клеток нуцеллуса и покровов. По мере развития зародышевого мешка клетки и ядра нуцеллуса и покровов разрушаются и исчезают. Например, у многих представителей семейства сложноцветных, имеющих тенуинуцеллятную семяпочку, ко времени полного сформирования зародышевого мешка от нуцеллуса не остается и следа и зародышевой мешок непосредственно граничит с эндотелием. В крассинуцеллятных семяпочках, в которых нуцеллус также частично или даже полностью разрушается, это происходит обычно после оплодотворения. У многих же растений по крайней мере эпидермис нуцеллуса сохраняется до созревания семени и входит в состав семенной кожуры.
После образования восьмиядерного зародышего мешка в микропилярной и халазальной частях его формируются по три клетки; в микропилярной части образуются три клетки яйцевого аппарата, имеющего непосредственное отношение к процессу оплодотворения. Одна из них является яйцеклеткой, т. е. женской гаметой, две сопутствующие ей — вспомогательными клетками, или синергидами. Иногда эти клетки могут быть очень сходны между собой, чаще они различаются размерами и расположением ядра и вакуоли. Для яйцеклетки характерно более крупное, чем у синергид, ядро, которое расположено обычно в апикальной части яйцеклетки, в то время как базальную ее часть занимает вакуоль. У синергид, как правило, расположение ядра и вакуоли обратное.
Форма и величина яйцеклеток у разных растений различны. Обычны грушевидные и цилиндрические яйцеклетки. Оболочка яйцеклетки тонкая, благоприятствующая обмену веществ.
Роль яйцеклетки ясна, так как именно она после оплодотворения дает начало новому организму — спорофиту; менее ясна роль синергид. Полагают, что синергиды играют важную роль в привлечении пыльцевых трубок в зародышевой мешок, а также в растворении оболочки пыльцевой трубки для освобождения ее содержимого. Существует мнение о том, что роль синергид заключается в обеспечении точного попадания спермиев в определенное место зародышевого мешка. Не выяснено пока лишь, как выполняется приписываемая синергидам роль в тех немногих случаях, когда они вовсе не образуются (например, у Plumbagella) или когда они разрушаются еще до оплодотворения.
В халазальной части зародышевого мешка нормального типа образуются три клетки с более или менее густым содержимым. Эти клетки называются антиподами. У многих растений число клеток антипод, первоначально равное трем, на более поздних стадиях развития зародышевого мешка значительно увеличивается. Например, у некоторых представителей семейства злаковых число антипод достигает 60, а у бамбуковых их еще больше — до 300. В таких случаях можно говорить об антиподальном комплексе клеток.
Клетки антиподального аппарата у различных видов растений варьируют как по размерам, так и по форме. В ряде случаев антиподальные клетки могут иметь не по одному, а по нескольку ядер, причем число ядер в одной клетке антипод может быть весьма значительным (например, у некоторых представителей семейства сложноцветных — до 40). Позднее в многоядерных антиподальных клетках отдельные ядра нередко сливаются, так что число их снова уменьшается, а сами ядра делаются более крупными, приобретая неправильную форму. Соответственно числу слившихся ядер в них возрастает и число хромосом: ядра становятся полиплоидными. Полиплоидность ядер антипод некоторых растений может осуществляться также путем эндомитоза.
У разных видов покрытосеменных растений антиподы проявляют различную жизнеспособность. В то время как у одних растений они разрушаются и исчезают еще до оплодотворения, у других — более или менее долго сохраняются и нередко присутствуют даже при образовании многоклеточного зародыша и эндосперма.
Некоторые авторы предполагают, что антиподы выделяют различные энзимы, способствующие разрушению прилегающих к ним клеток семяпочки; освобождающиеся в результате разрушения этих клеток вещества абсорбируются антиподами, а затем ассимилируются развивающимся зародышевым мешком. Оставшиеся в восьмиядерном зародышевом мешке после образования яйцевого и антиггодального аппаратов два ядра (по одному у каждого полюса) называются полярными ядрами; они приближаются друг к другу и сливаются, образуя ядро центральной клетки, называемое иначе вторичным, или центральным, ядром зародышевого мешка. У некоторых растений полярные ядра лишь приближаются друг к другу, не сливаясь до оплодотворения. Число ядер, входящих в состав вторичного ядра, может быть различным в зависимости от типа развития зародышевого мешка. Из центральной клетки после ее оплодотворения образуется эндосперм — особая ткань, содержащая запасные питательные вещества, используемые зародышем в процессе его развития.
Ядро центральной клетки обычно располагается непосредственно под яйцеклеткой, у некоторых растений оно лежит в центре зародышевого мешка или даже в халазальной его части.
В цитоплазме зародышевого мешка, а также в цитоплазме яйцеклетки обнаруживаются различные включения: митохондрии, хондриосомы, пластиды, крахмал, жир и т. д.
Процесс оплодотворения, характеризующийся слиянием половых клеток — мужской и женской, — непременное условие нормального полового размножения, или амфимиксиса. Биологическое значение этого способа размножения заключается в том, что благодаря двойственной наследственности (материнской и отцовской) возникающие организмы оказываются более жизнеспособными и приспособленными к варьирующим условиям окружающей среды, чем при бесполом размножении.
Для того чтобы оплодотворение могло произойти, прежде всего необходимо опыление, т. е. пыльца из пыльников должна попасть на рыльце пестиков и прорасти на них. В процессе эволюции у растений выработались различные приспособления для опыления.
У одних растений осуществляется перенос пыльцы с тычинок цветка одного растения на рыльце пестика цветка другого. Такое опыление называется перекрестным (аллогамия, или ксеногамия). У других пыльца переносится на рыльце того же самого цветка. Такое опыление называется самоопылением (автогамия).
В некоторых случаях самоопыляющиеся цветки вообще не раскрываются. Пыльца в таких цветках не высыпается на рыльце, а прорастает в пыльниках, пыльцевые трубки достигают рыльца и растут дальше. Такое скрытное опыление называется клейстогамией. Клейстогамные цветки известны у фиалок, кислицы, арахиса и ряда других растений.
В тканях пестика пыльцевые трубки растут или эктотропно, т. е. по каналам или по поверхности проводниковой ткани, или эндотропно, т. е. внутри проводниковой ткани или по межклетникам других тканей, раздвигая рыхло соединенные клетки. Дойдя до семяпочки, пыльцевая трубка проникает в зародышевый мешок либо непосредственно через микропиле, либо попадает в него окольными путями. Первый тип прохождения пыльцевой трубки, встречающийся наиболее часто, называется порогамией, или акрогамией. Второй тип — апорогамия — встречается значительно реже. Частный случай апорогамии — проникновение пыльцевой трубки через халазу семяпочки — называется халазогамией, или базигамией. Между порогамией и халазогамией имеется промежуточный тип, известный под названием мезогамии.
При прорастании пыльцы, после того как пыльцевая трубка достигнет некоторой длины и проникнет в ткань верхней части пестика, в нее начинает переходить содержимое пыльцевого зерна. Наибольшее количество цитоплазмы концентрируется в кончике пыльцевой трубки, сюда же постепенно перемещаются вегетативное ядро и генеративная клетка или вегетативное ядро и спермии.
Достигнув зародышевого мешка, пыльцевая трубка изливает свое содержимое чаще всего в одну из синергид, которая при этом разрушается. Иногда при проникновении пыльцевой трубки в зародышевый мешок затрагиваются обе синергиды, вследствие чего они разрушаются. В редких случаях, например у некоторых орхидей, обе синергиды могут не затрагиваться пыльцевой трубкой. Хотя в столбик пестика обычно входит много пыльцевых трубок, большинство из них по мере своего продвижения к зародышевому мешку прекращают рост и не доходят до семяпочки и зародышевого мешка. Рост пыльцевых трубок может остановиться на разных расстояниях от зародышевого мешка, при этом на концах пыльцевых трубок обычно образуются булавовидные вздутия, наполненные густым содержимым, вызывающим закупорку пыльцевых трубок.
После своего освобождения из пыльцевых трубок спермин направляются один к яйцеклетке, другой к ядру центральной клетки и сливаются с ними. Этот процесс, открытый в 1898 г. русским ученым С. Г. Навашиным, получил название двойного оплодотворения.
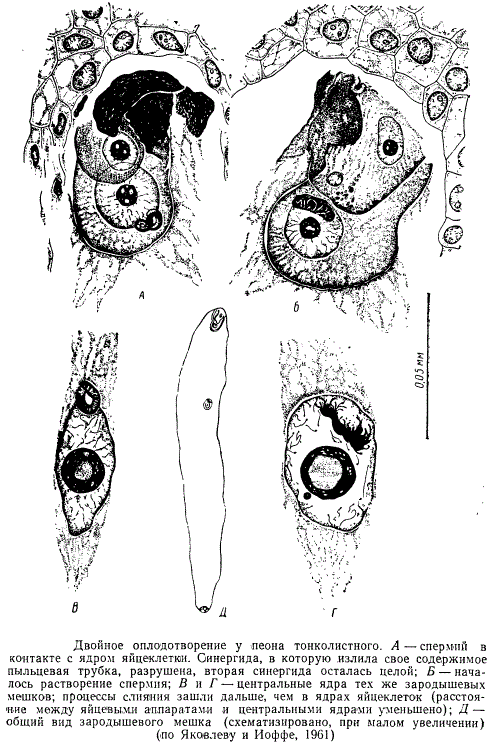
По выходе спермиев из пыльцевой трубки последняя вместе с остатками своего содержимого (цитоплазмой, вегетативным ядром, если оно дошло до зародышевого мешка, и разными включениями) ассимилируется зародышевым мешком, который после оплодотворения яйцеклетки и центральной клетки начинает интенсивно расти.
У некоторых растений, например у злаковых, полярные ядра до самого оплодотворения остаются неслившимися; в таких случаях слияние их друг с другом происходит одновременно со слиянием их со спермием (пшеница) или несколько позднее — во время профазы первого деления зачатка эндосперма (кукуруза). Ядра спермиев у разных видов растений имеют различную форму: они могут быть округлыми, спиральными, червеобразными и т. д.
У большинства покрытосеменных растений промежуток времени между опылением и оплодотворением очень непродолжителен и измеряется днями, часами и даже минутами. У большинства голосеменных растений этот период значительно более продолжителен и измеряется месяцами. Однако есть покрытосеменные, например некоторые представители однопокровных, у которых этот период так же продолжителен, как и у большинства голосеменных: так, у некоторых видов дуба с двулетним периодом развития семян он равен 12 и даже 14 месяцам; у представителей семейства березовых он колеблется от 1 до 4 месяцев. У большинства же других покрытосеменных продолжительность периода от опадения до оплодотворения значительно короче. У груши и яблони оплодотворение происходит через 5 дней после опыления 3 разных видов лилий промежуток времени между опылением и оплодотворением колеблется от 2 до 7 суток. Наименьший промежуток времени между опылением и оплодотворением обнаружен у некоторых сложноцветных с колебаниями от 1,5 часов до 10 минут. Например, у кок-сагыза оплодотворение происходит через 15—20 минут после опыления.
Внешние условия, особенно изменения температуры и влажности, влияют на скорость роста пыльцевых трубок, оплодотворения, а также на развитие зародыша и эндосперма. Повышение температуры и понижение влажности до известного предела обычно ускоряют эти процессы, понижение же температуры и повышение влажности замедляет их и даже останавливает. Так же действуют слишком высокая температура и слишком низкая влажность.
В зародышевый мешок, как правило, входит одна пыльцевая трубка, но нередки случаи, когда в одном и том же зародышевом мешке можно обнаружить несколько пар спермиев. Например, в зародышевом мешке грецкого ореха, скерды, кок-сагыза, подсолнечника и других растений некоторые исследователи наблюдали от 2 до 10 пар спермиев.
Яйцеклетка и центральная клетка оплодотворяются одной парой спермиев (моноспермия), но при вхождении в зародышевый мешок нескольких пыльцевых трубок в редких случаях яйцеклетка может оплодотвориться двумя (диспермия) и даже несколькими (полиспермия) спермиями.
В исключительных случаях, помимо яйцеклетки и центральной клетки, могут быть оплодотворены и другие клетки зародышевого мешка — синергиды и антиподы.
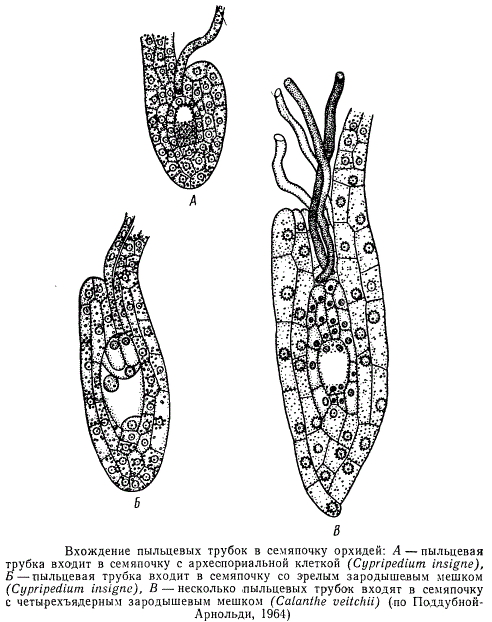
Обычно к моменту оплодотворения в завязях имеются семяпочки с вполне сформировавшимся зародышевым мешком. Однако у некоторых растений, например у цитрусовых, пыльцевые трубки подходят к семяпочке в тот момент, когда зародышевый мешок в ней еще не сформирован и находится на более или менее ранних стадиях развития. Известны даже такие растения, у которых к моменту опыления семяпочки в завязи отсутствуют и закладываются только после опыления. Наиболее показательным примером в данном отношении является семейство орхидных. У некоторых его представителей до опыления зародышевый мешок не только не развивается, но даже не образуются сами семяпочки. Если опыления не произойдет, семяпочки так и не разовьются. На рисунке, изображающем семяпочку Cypripedium insigne, видно, что пыльцевая трубка находится в микропиле, в то время как в семяпочке находится еще только археспориальная клетка.
После опыления и оплодотворения происходит усиленный приток питательных и физиологически активных веществ к завязи, что стимулирует ее дальнейшее развитие и превращение в плод. Нормальное течение процессов опыления и оплодотворения точно так же, как и процесс развития пыльцы и зародышевого мешка, может быть нарушено под воздействием различных факторов. Неблагоприятная погода, недостаток питания и опылителей, слишком высокая или низкая температура, высокие дозы различных химикалиев и рентгеновских лучей, отдаленная гибридизация, полиплоидия, физиологическая и генетическая несовместимость и т. д. могут нарушить нормальный ход этих процессов. Эти нарушения обычно выражаются в замедленном прорастании пыльцы, в замедленном росте пыльцевых трубок и даже полной его остановке, в увеличении промежутка времени между опылением и оплодотворением, в потере жизнеспособности половыми клетками, что приводит к частичной или полной стерильности.