Сосудисто-волокнистые пучки и механические ткани во всех органах погружены в основную паренхиму.
Паренхима может быть и довольно плотной и очень рыхлой тканью. Оболочки паренхимных клеток чаще тонкие, но могут достигать и весьма значительной толщины, причем клеточная полость таких клеток значительно уменьшается. Оболочки клеток основной паренхимы либо остаются на все время существования целлюлозными, как, например, в коре ряда растений, либо одревесневают, как это происходит в сердцевине древесных и многих травянистых растений.
В осевых органах основная паренхима сгруппирована в трех достаточно отчетливо дифференцирующихся участках, составляя сердцевину, сердцевинные лучи и кору. В листьях в соответствии с характером и особенностями развития также можно наметить по существу те же категории паренхимных тканей. Но вследствие особой функции листа эти ткани расположены иначе и своеобразно видоизменены.
Сердцевина состоит из клеток, обычно незначительно вытянутых по длине органа. Клетки сердцевины иногда сильно различаются по размерам. Например, участки сердцевины, непосредственно примыкающие к центральному цилиндру, почти всегда состоят из клеток, более мелких, чем ее центральная часть.
Зона сердцевины, ближайшая к центральному цилиндру, называется перимедуллярной зоной. Оболочки клеток этой зоны обычно несколько толще, чем в центральных участках сердцевины нормальной структуры.
Вначале все клетки сердцевины живые, позже некоторые клетки не только отмирают, но в ряде случаев разрушаются, образуя полость в центре стебля, причем клетки перимедуллярной зоны остаются живыми и деятельными дольше всех. У некоторых растений, например у бука, клетки сердцевины остаются жизнедеятельными около 40 лет. Живые клетки нередко перемежаются с мертвыми. Иногда расположение живых и мертвых, крупных и мелких клеток подчиняется определенной закономерности, причем некоторые семейства, роды и виды растений имеют только им свойственные комбинации в сочетаниях клеток.
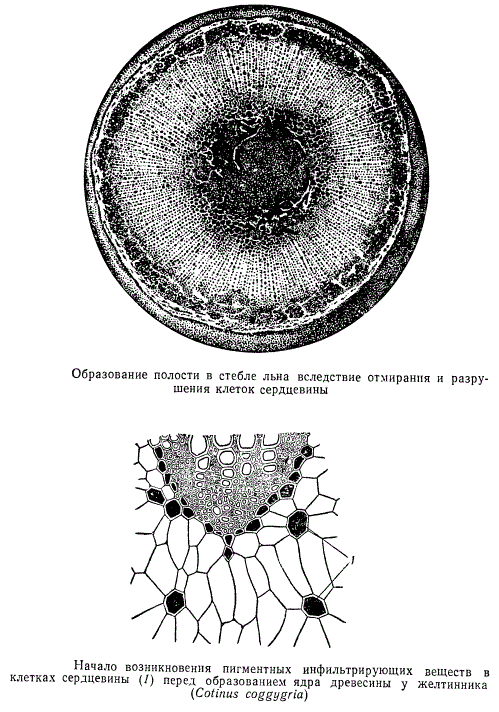
В сердцевине часто отлагаются дубильные вещества, образуются различные пигменты внепластидного происхождения, имеющие отношение к формированию ядра древесины. У ряда растений в сердцевине образуются вместилища для отложения продуктов секреторной деятельности, а также каналы, содержащие смолу, каучук и другие вещества.
Вообще сердцевина, вопреки предполагаемой в ней с первого взгляда простоте структуры, весьма сложная и многосторонняя по строению и функциям ткань. Она может состоять из округлых, угловатых или вытянутых в том или другом направлениях клеток. В одних случаях клетки сердцевины по форме и по содержимому однородны, в других — исключительно разнородны. В такой же степени, если не более, разнообразна другая паренхимная ткань осевого органа — первичная кора. В сердцевине, как правило, нет механических элементов, исключая каменистые клетки, но в коре, в первую очередь, часто развивается очень характерная механическая ткань — колленхима, структура которой была описана выше. Кроме основной защитной роли, первичная кора, подобно сердцевине, может играть и роль места отложения запасных веществ. В первичной коре легче и потому чаще, чем в сердцевине, образуются хлоропласты, и она может до некоторой степени участвовать в фотосинтезирующей деятельности растительного организма. Эта деятельность наиболее наглядна в зеленых побегах, лишенных листьев, а также в так называемых филлокладиях (например, у кактусов, иглицы и пр.).
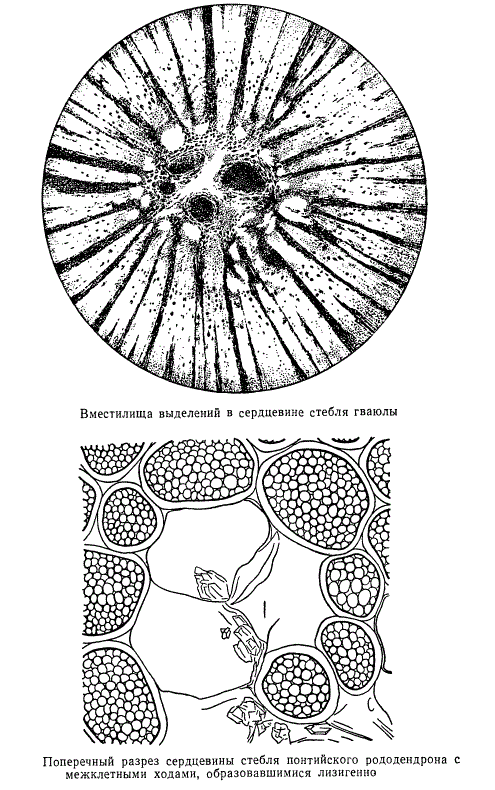
У некоторых растений кора построена очень рыхло, значительно более рыхло, чем сердцевина, с обширными воздушными ходами. Особенно мощно система воздухоносных полостей у растений, произрастающих во влажных местах и в воде, развита в коре и сердцевине.
В общем первичная кора сходна с сердцевиной не только по своей структуре, но и по происхождению.
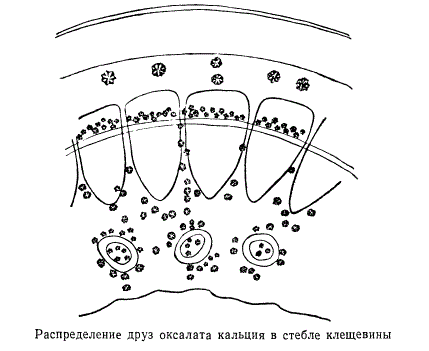
В состав вторичной коры, кроме основной паренхимы, входят различные анатомические элементы лубяной части сосудисто-волокнистых пучков. Паренхима вторичной коры образует преимущественно коровую часть сердцевинных лучей. У некоторых растений, как, например, у виноградной лозы, здесь наблюдается своеобразное расположение кристаллов оксалата кальция. Клетки, содержащие кристалические отложения, представляют собой подобие барьера, отделяющего сердцевинный луч от лубяной части центрального цилиндра осевого органа. Кристаллоносные клетки обычно располагаются в определенном порядке, будучи, по-видимому, тесно связанными с какими-то закономерными функциями растительного организма. Пример отчетливо выраженной закономерности в расположении кристаллоносных клеток изображен на рисунке. Здесь показано распределение друз оксалата кальция в коре, лубяной части проводящих пучков, около камбия, в сердцевине и сердцевинных лучах.
Сердцевинные лучи в осевых органах первичного строения состоят из основной паренхимы, тождественной по своей структуре с паренхимой первичной коры и сердцевины.
У однодольных растений сосудисто-волокнистые пучки обычно рассеяны по всему массиву основной паренхимы, как, например, в стеблях злаков, не имеющих полости внутри, в стеблях лилейных, пальм и пр. При такой организации осевого органа сердцевинных лучей быть не может.
У двудольных (особенно древесных), а также у голосеменных в связи с образованием вторичных утолщений структура сердцевинных лучей в большинстве случаев сильно меняется. У травянистых же двудольных, у которых сосудисто-волокнистые пучки преимущественно расположены обособленно, структура серцевинных лучей по сравнению с их первичным строением меняется незначительно.
На поперечных разрезах древесных побегов сердцевинные лучи часто имеют вид узких длинных полос, идущих по радиусам от сердцевины к коре. Внимательно рассматривая срезы древесины, можно убедиться, что ширина сердцевинных лучей у одного и того же растения либо на всем протяжении одинакова, либо одни сердцевинные лучи шире других. На продольных тангентальных разрезах оси побегов видно, что и высота сердцевинных лучей различна. У одних растений сердцевинные лучи на тангентальных разрезах имеют вид довольно высоких полос, у других большинство из них напоминают небольшие узкие линзы, как бы вкрапленные в срез.
По отношению к массиву древесины площадь, занимаемая всеми сердцевинными лучами каждого дерева, незначительна. На поперечных разрезах одревесневшего побега легко обнаружить, что не все они доходят до сердцевины, некоторые лучи начинаются в толще древесины на различных расстояниях от поверхности органа. Это вторичные лучи. Лучи, идущие от сердцевины, являются первичными. Однако во многих случаях и первичные лучи в начале развития органа не доходят до сердцевины, а упираются в протоксилему, как почти всегда бывает в корнях. На основании этого некоторые исследователи находят название «сердцевинные лучи» нерациональным, предлагая взамен этого термин «лучи». Мы все же для удобства будем придерживаться общепринятой номенклатуры.
Вторичные сердцевинные лучи, в отличие от первичных, возникают из камбия. Среди клеток камбия, откладывающих веретенообразные элементы, дифференцируются одна или несколько клеток, образующих паренхимные элементы — начальные клетки сердцевинных лучей. Работа таких образовательных клеток однообразно продолжается до конца существования растения. Начальные клетки сердцевинных лучей возникают из камбия в сравнительно небольших количествах каждый год.
Часть сердцевинного луча, проходящего по коре, отличается от ксилемной части его тем, что стенки клеток в ксилемной части одревесневшие, а во флоэмной — целлюлозные. Особенно отчетливо это различие выражено у древесных растений. Часть сердцевинного луча, проходящая в коре, отличается от его древесинной части еще тем, что в древесине луч почти совершенно не расширяется, в коре же некоторые, особенно первичные, сердцевинные лучи к периферии становятся значительно шире, чем в участке около камбия. Такое расширение сердцевинных лучей в коре можно видеть, например, у липы, у виноградной лозы. У лиан, подобно винограду, сердцевинные лучи расширяются даже в древесинной части. В корнях расширение сердцевинных лучей встречается чаще, чем в стеблях.

Сердцевинные лучи могут состоять из одного или нескольких рядов клеток, как хорошо видно на поперечных разрезах. Лучи, состоящие только из одного ряда клеток, называются однорядными, из двух рядов — двурядными, а более чем из двух рядов — многорядными. Узкие однорядные сердцевинные лучи на тангентальных разрезах древесных стволов представляют собой цепочку паренхимных клеток, расположенных друг за другом в вертикальном направлении. Чаще всего однорядные лучи образуются в древесине хвойных. Многорядные сердцевинные лучи характерны в основном для травянистых растений, а также для лиан. Среди древесных двудольных растений значительной шириной сердцевинных лучей отличается дуб. У одних двудольных растений в сердцевинных лучах краевые клетки значительно короче срединных, ничем в других отношениях не отличаясь от последних. Это так называемые гомогенные лучи. У других же растений краевые клетки очень высокие и резко отличаются от срединных клеток. При такой структуре сердцевинного луча краевые клетки называются стоячими, срединные — лежачими, а сам луч называется гетерогенным.
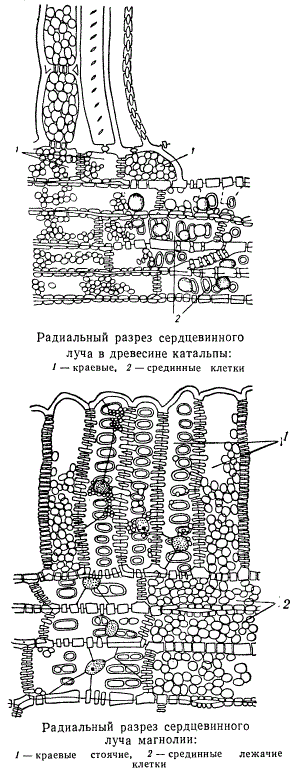
У двудольных растений все клетки сердцевинных лучей живые и в них периодически накапливаются различные запасные вещества. Различия клеток в разных частях луча часто зависят от того, что одни из них соединяются порами с сосудами, примыкающими к сердцевинному лучу, другие — нет. В клетках луча, снабженных порами, обращенными к сосуду, запасные вещества изменяются более энергично, чем в клетках луча, соединяющихся порами с какими-либо другими элементами древесины.
На поперечных разрезах, проходящих через узкий сердцевинный луч двудольного, зависимость состояния содержимого клеток сердцевинного луча от того, соединяется ли с сосудом клетка непосредственно или через другие клетки, отчетливо видна. На рисунке изображен поперечный разрез участка древесины грецкого ореха, проходящий через группу, состоящую из трех сосудов, которые огибает однорядный сердцевинный луч.
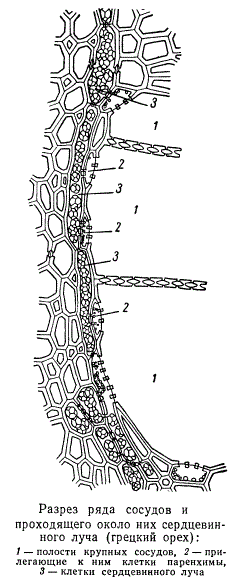
Одна из клеток сердцевинного луча входит в контакт с одним из сосудов. Содержимое этой клетки отличается от содержимого других клеток сердцевинного луча, входящих в контакт с сосудом лишь через посредство паренхимы, расположенной между сердцевинным лучом и сосудом. Та же самая зависимость содержимого клеток сердцевинного луча от типа связи их с сосудом может быть обнаружена и на радиальных продольных разрезах.
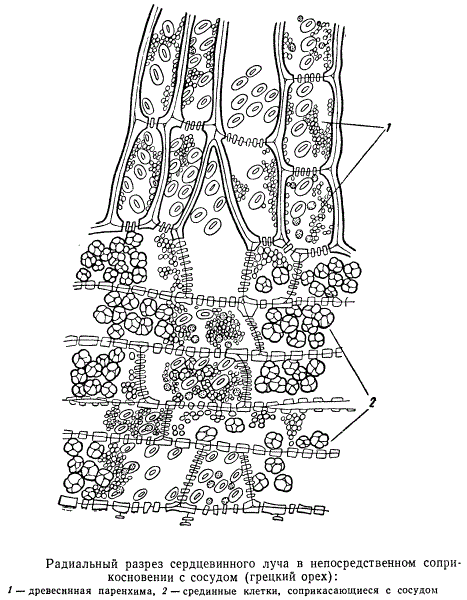
У многих хвойных клетки, расположенные по краям сердцевинного луча в его древесинной части, во вполне оформившемся состоянии не имеют живого содержимого и сообщаются друг с другом, а также с соседними трахеидами при помощи окаймленных пор. Центральные, живые клетки сердцевинных лучей с деятельными пластидами соединяются со всеми соприкасающимися с ними анатомическими элементами простыми порами.
Многорядные сердцевинные лучи принципиально имеют ту же структуру, что и однорядные, с такой же дифференциацией клеток и их содержимого. Часто для многорядных сердцевинных лучей характерны воздушные ходы, пронизывающие лучи в радиальном направлении. В коре сердцевинные лучи оканчиваются там, где есть чечевички. Несомненно, что воздушные ходы осуществляют аэрацию глубоких слоев древесины и что отдушинами для этого служат чечевички.
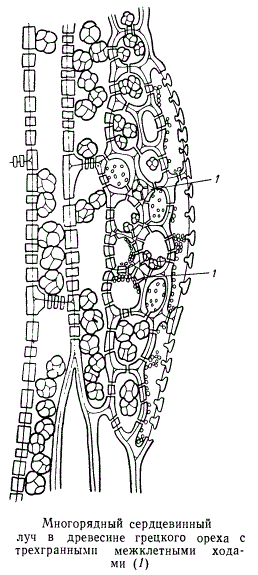
Сердцевинные лучи почти всегда содержат в своих клетках запасные вещества. Во время распускания листьев, цветения и особенно плодоношения отложенные в клетках сердцевинных лучей энергопластические вещества в значительной мере мобилизуются. Если следить за состоянием отложенных в клетках сердцевинных лучей веществ непрерывно в течение года, можно установить передвижение этих веществ по сердцевинным лучам из коры в древесину и обратно. Сердцевинные лучи служат путями передвижения пластических веществ по древесине в радиальном направлении. Разумеется, по сердцевинным лучам может передвигаться и вода. У хвойных краевые клетки сердцевинных лучей по своей структуре — вполне водопроводящие элементы, отличающиеся от типичных трахеид лишь формой и размерами.
Клетки сердцевинных лучей двудольных очень долго остаются живыми. Например, у граба в сердцевинных лучах даже на глубине 107-го годичного кольца, считая от границы древесины с корой, клетки были живыми; у бука живые клетки в сердцевинных лучах найдены в 98-м годичном слое; у яблони — на уровне 24-го годичного слоя древесины.
Организация системы межклеточных ходов, пронизывающих древесину, показывает, что аэрация массы древесины, как бы она ни была обширна, осуществляется в стволах древесных пород достаточно хорошо. Межклетные ходы могут достигать значительной длины.