В каждом семени образуется в основном один зародыш. В некоторых случаях в семени может развиться несколько зародышей. Это явление получило название многозародышевости, или полиэмбрионии.
Зародыши возникают не только из яйцеклетки, но и почти из всех клеток женского гаметофита — из синергид и из антипод. В ряде случаев своеобразное развитие зиготы приводит к созданию многоклеточного образования, которое впоследствии или само распадается на несколько проэмбрио, или на основе его развивается несколько зародышей. Дополнительные зародыши образуются также при почковании самого зародыша или его подвеска. Иногда несколько зародышей возникает за счет формирования их из разных зародышевых мешков, развивающихся в одном нуцеллусе.
Зародыши могут появиться также из соматических клеток нуцеллуса и интегумента. Иногда несколько зародышей в семени возникает в результате развития двух нуцеллусов в одной семяпочке, каждый из которых имеет свой зародышевый мешок (тератологический случай).
В случаях полиэмбрионии одни зародыши образуются в результате оплодотворения, другие — без оплодотворения (апомиктично). Поэтому зародыши, развивающиеся в одном семени, могут иметь гаплоидные, диплоидные или полиплоидные числа хромосом.
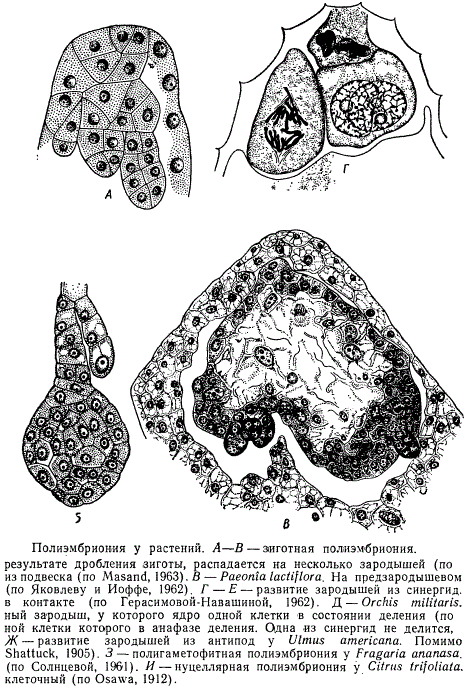
Полиэмбриония не всегда приводит к образованию зародышей, одинаково способных к прорастанию. Чаще всего большинство зародышей погибает на ранних стадиях развития и к моменту созревания сохраняются лишь некоторые из них. Если же в готовом семени находятся несколько зародышей, они обычно характеризуются разной степенью дифференциации. Вследствие недоразвития и, возможно, недостаточного питания не все зародыши, развившиеся в семени, дают проростки.
Образование плода начинается обычно после оплодотворения, развитие же семени идет параллельно с развитием плода; если семя не образуется, то обычно не развивается и плод.
У некоторых растений, однако, плоды развиваются, несмотря на то, что семена не образуются из-за дегенерации или недоразвития семяпочек. Такие плоды часто отличаются от нормально развившихся плодов меньшими размерами и рядом других качеств. Явление образования плодов без оплодотворения, лишенных семян, называется партенокарпией. При этом в одних случаях происходит опыление и тогда осуществляется стимулятивная, или индуцированная, партенокарпия, в других — отсутствует и опыление и осуществляется автономная (неиндуцированная), или вегетативная, партенокарпия.
Партенокарпия может быть случайной и постоянной. Она обнаружена как у дикорастущих, так и культурных видов покрытосеменных растений. Среди культурных растений она хорошо выражена у груши, яблони, винограда, мандарина, лимона, банана, апельсина, ананаса, инжира, тыквы, огурцов, томата и т. д.
Партенокарпия имеет большое практическое значение, так как плоды, лишенные семян, отличаются чрезвычайной мясистостью, сочностью и хорошими вкусовыми качествами. Что касается развития пыльцы и зародышевого мешка у партенокарпических видов, то при этом возможно как нормальное их образование, так и нарушение обычного процесса их формирования и дегенерация на разных стадиях развития.
Наряду с развитием бессемянных плодов без оплодотворения у некоторых растений могут развиваться семена без зародыша. Это явление называется партеноспермией. Особенно часто партеноспермия обнаруживается у хвойных, в частности у лиственницы.
Наряду с нормальным половым размножением (амфимиксисом), вследствие которого организмы возникают путем слияния мужской и женской гамет, у высших растений встречается и другой способ размножения, называемый апомиксисом. Апомиксис — это такой способ размножения, при котором развитие нового организма происходит без слияния половых клеток, но и при этом сохраняются морфологические структуры, связанные с половым размножением.
Несмотря на то что при апомиксисе не происходит объединения гамет, в ряде случаев необходимо воздействие мужского начала на развитие семени. Апомиктическое развитие может осуществляться также и без всякого воздействия мужских элементов. Такие апомикты называются автономными. Если апомиктическое развитие вызывается (стимулируется) отцовским началом, то такие апомикты называются индуцированными, стимулятивными.
Влияние мужских элементов может быть более или менее глубоким. В одних случаях для апомиктического развития достаточно одного лишь опыления — присутствия пыльцы на рыльце пестика, которая даже не прорастает. В других — необходимо прорастание пыльцевых зерен, воздействие со стороны растущей пыльцевой трубки. Для некоторых апомиктов и такого воздействия оказывается недостаточно. Для их развития необходимо, чтобы пыльцевая трубка проникла в зародышевый мешок, и даже чтобы один из спермиев слился с центральным ядром зародышевого мешка. Такое апомиктическое развитие называется псевдогамным. Иногда термином «псевдогамия» обозначают все случаи индуцированного апомиксиса.
В настоящее время существует ряд классификаций апомиксиса. Здесь дается классификация В. А. Поддубной-Арнольди (1964).
- Партеногенез — образование зародыша из неоплодотворенной яйцеклетки с нередуцированным числом хромосом в случаях нередуцированного партеногенеза и с редуцированным в случаях редуцированного партеногенеза:
а) нередуцированный партеногенез, связанный с псевдогамией, т. е. зависящий от воздействия пыльцевой трубки;
б) нередуцированный партеногенез, не связанный с псевдогамией, т. е. не зависящий от воздействия пыльцевой трубки;
в) редуцированный женский партеногенез, или гиногенез, связанный с псевдогамией, — партеногенетическое образование сходной с матерью особи с материнским набором хромосом после опыления своей или чужой пыльцой при отсутствии слияния женской и мужской гамет и дегенерации последней, но при наличии мейоза;
г) редуцированный мужской партеногенез, или андрогенез, связанный с псевдогамией, — партеногенетическое образование сходной с отцом особи с отцовским набором хромосом после опыления своей или чужой пыльцой при отсутствии оплодотворения и при дегенерации ядра яйцеклетки, но при наличии мейоза.
Некоторые исследователи приравнивают мужской партеногенез к мерогонии, т. е. образованию зародыша после оплодотворения спермиями безъядерных участков яйцеклетки — явлению, впервые описанному у морских ежей.
- Апогаметия — возникновение зародыша не из яйцеклетки, а из других клеток женского гаметофита с нередуцированным числом хромосом в случаях нередуцированной апогаметии и с редуцированным в случаях редуцированной апогаметии:
а) нередуцированная апогаметия — образование зародыша из клеток синергид и антипод с нередуцированным числом хромосом;
б) редуцированная апогаметия — образование зародыша из клеток синергид и антипод с редуцированным числом хромосом.
- Апоспория — развитие гаметофита не из споры, а из гаплоидной или диплоидной клеток спорофита. У покрытосеменных растений до сих пор наблюдалась лишь апоспория, связанная с диплоидным числом хромосом. Некоторые исследователи различают у покрытосеменных две формы апоспории:
а) эвапоспория — развитие зародышевого мешка из чисто соматических клеток, из клеток нуцеллуса и покровов семяпочки;
б) псевдоапоспория, или соматическая апоспория, — развитие зародышевого мешка из соматических клеток нуцеллуса семяпочки, но родственных археспориальным клеткам.
- Нуцеллярная эмбриония — образование зародышей вне зародышевого мешка из клеток нуцеллуса семяпочки:
а) индуцированная нуцеллярная эмбриония, связанная с оплодотворением или, по крайней мере, с опылением;
б) автономная неиндуцированная нуцеллярная эмбриония, не связанная ни с оплодотворением, ни с опылением.
- Интегументальная эмбриония — образование зародышей вне зародышевого мешка из клеток покровов семяпочек:
а) индуцированная интегументальная эмбриония, связанная с оплодотворением или, по крайней мере, с опылением;
б) автономная неиндуцированная интегументальная эмбриония, не связанная ни с опылением, ни с оплодотворением.
Апомиктический способ размножения у многих растений является наследственным, постоянным, у других растений апомиксис ненаследственный и случайный.
Внутри одного и того же вида, или биотипа, наряду с апомиктическим способом размножения может встречаться и нормальное половое размножение. Такой апомиксис называют частичным. Апомиксис называют полным, когда все биотипы этого вида размножаются только апомиктично.
Апомиктичный способ размножения в определенных условиях имеет большие преимущества по сравнению с амфимиксисом. Неблагоприятные внешние условия (недостаток насекомых, плохая погода), затрудняющие нормальное опыление и оплодотворение, не мешают многим апомиктичным видам давать большое количество зрелых семян. Даже наоборот, крайние условия существования провоцируют у растений, размножающихся половым путем, появление апомиктичных форм. Этим и объясняется то обстоятельство, что местом возникновения апомиктичных форм становятся границы ареала исходного вида. Переход на апомиктичный путь размножения способствует закреплению тех признаков и свойств растения, которые у него были к моменту
появления апомиксиса. Постоянное однородное потомство у апомиктов — характерная черта этого способа размножения. Если внешняя среда устойчива и изменяется очень медленно, то появившиеся апомикты процветают, несмотря на их малую пластичность. При быстром и резком изменении среды апомикты не успевают приспособиться к этим условиям и в конце концов погибают. В тех случаях, когда апомиксис не полный, не облигатный, растениям удается сочетать преимущества полового и апомиктичного способа размножения и колеблющиеся условия среды не приводят к исчезновению апомиктов вследствие их малой пластичности. При таких условиях обеспечивается не только длительное существование, но и процветание вида.
С этой точки зрения и следует рассматривать эволюционное значение апомиксиса.
До последнего времени относительно значения апомиксиса в эволюции существовали разные мнения. Одно из них состояло в том, что апомиктичное размножение, будучи приспособлением к сильноспециализированным, устойчивым условиям среды, заводит формы в тупики эволюции, из которых они не могут выйти на основную, генеральную линию эволюции. Другие считают, что апомиктичный способ размножения у покрытосеменных совершеннее, чем половой, и в будущем должен его вытеснить.
Наконец, существует и третья точка зрения, согласно которой апомиксис рассматривается как особая линия прогрессивной эволюции, не являющаяся эволюционным тупиком. В сочетании с половым размножением (и только при этом) апомиксис способен обеспечить длительное сохранение вида при резких изменениях внешней среды и даже служить основой для эволюции. Однако апомиктическое размножение не может полностью заменить половое размножение, так как при полном вытеснении полового размножения апомиктическое размножение при быстрых и резких изменениях внешней среды ведет к вырождению и к гибели вида и действительно превращается в эволюционный тупик.