В 1927 г. в Берлине состоялся V Международный генетический конгресс.
Доклад, прочитанный на этом конгрессе американским ученым Г. Д. Мёллером о возможности получения мутаций под влиянием рентгеновских лучей у Drosophila melanogaster, вызвал сенсацию. Это открытие послужило началом новой эры в генетике. В течение последующих 15—20 лет было показано, что у всех организмов, от вирусов до млекопитающих, под влиянием рентгеновских лучей возникают мутации и что все ионизирующие излучения также являются мутагенными. В 1946 г. д-р Мёллер был удостоен Нобелевской премии.
За двадцать восемь лет своего существования радиационная генетика превратилась в обширную и плодотворную отрасль исследования. С точки зрения задач настоящей книги нас интересует главным образом ответ на два вопроса:
- какие типы мутаций вызывают рентгеновские лучи;
- какова частота возникновения этих мутаций при определенной дозе рентгеновских лучей.
Рассмотрим эти вопросы поочередно.
Типы мутаций, вызываемых рентгеновскими лучами
Рентгеновские лучи оказывают на клеточное ядро двоякое действие. Во-первых, они вызывают генные мутации, которые могут быть доминантными или рецессивными; рецессивные мутации возникают значительно чаще.
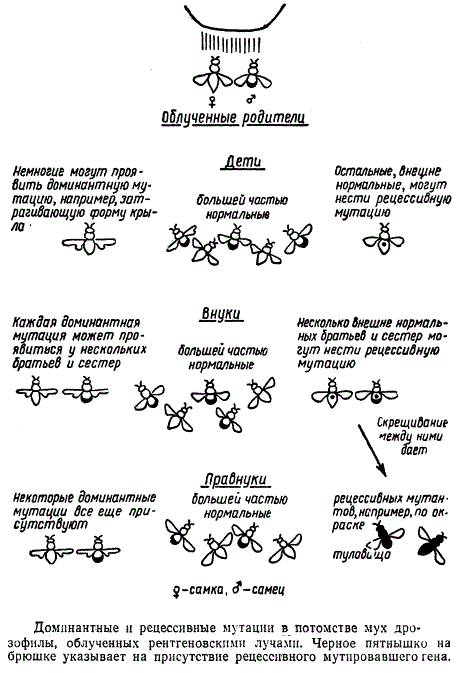
Рисунок иллюстрирует, каким образом обнаруживается генная мутация в потомстве мух дрозофил, подвергавшихся воздействию рентгеновских лучей.
Вспомним, что доминантная мутация, подобно мутации «курчавости» у кур, проявляется у каждой особи, получившей мутировавший ген от одного из родителей. Появившиеся под влиянием рентгеновских лучей доминантные мутации будут, следовательно, обнаружены у детей облученных родителей.
Рецессивная мутация, подобно мутации рыжей окраски у спаниеля, проявляется только у особей, получивших одинаковые мутировавшие гены от обоих родителей. Крайне маловероятно, чтобы даже после облучения обоих родителей сперматозоид и яйцеклетка, которую он оплодотворяет, могли нести одинаковые мутировавшие гены. Вследствие этого вызванные рентгеновскими лучами рецессивные мутации не проявятся у детей облученных родителей, но когда у потомка, несущего новый рецессивный ген, в свою очередь, появятся дети, половина из них унаследует новый рецессивный ген. Среди внуков облученного поколения появится несколько братьев и сестер, несущих, но не проявляющих один и тот же новый рецессивный ген. Если такие особи скрестятся между собой, некоторые правнуки могут получить этот ген от обоих родителей и его действие проявится. Следовательно, если у определенного вида происходят скрещивания между братьями и сестрами, рецессивные мутации смогут впервые проявиться лишь у правнуков облученного поколения. Не все рецессивные гены, конечно, смогут обнаружиться так скоро. Для выявления некоторых из них потребуется значительно больше времени. У человека браки между братьями и сестрами запрещены и даже браки между кузенами редки, поэтому должно пройти обычно много поколений, прежде чем вызванные рентгеновскими лучами рецессивные мутации проявят свое действие.
Мутации, полученные под влиянием рентгеновских лучей, подобно другим мутациям, обусловлены процессом случайного характера, и изменения, которые они вызывают, обычно вредны для организма.
Доминантные мутации, приводящие к появлению видимых ненормальностей среди детей облученных индивидуумов, очень редки у дрозофилы и мышей. Рецессивные мутации встречаются гораздо чаще. Большинство видимых мутантных особей, появляющихся в потомстве облученных дрозофил, слабее, менее плодовиты и менее долговечны, чем нормальные мухи.
Значительно более многочисленными по сравнению с мутациями, обладающими видимым действием, являются мутации, уменьшающие выносливость организма, продолжительность жизни и плодовитость, но не меняющие внешний вид особи. Часто вредное действие оказывается слабым, и его легко можно не заметить, однако соответствующая методика скрещиваний и учета показала, что рецессивные мутации, обладающие слегка вредным действием (и отсутствием видимого), представляют собой самую обширную категорию генных мутаций, вызываемых рентгеновскими лучами у дрозофилы. Быть может, не столь многочисленны, но все же значительно более часто, чем видимые мутации, встречаются рецессивные летали, убивающие развивающийся организм. Многие правнуки облученных мух погибают в процессе развития из-за наличия рецессивных леталей, унаследованных ими от обоих родителей.
Представляется вероятным, что и у человека рентгеновские лучи будут вызывать больше рецессивных, чем доминантных мутаций, и среди обеих категорий будут преобладать мутации с вредным или летальным действием.
Исключение из этого правила, согласно которому рецессивные мутации впервые обнаруживаются у правнуков облученного поколения, составляют рецессивные мутации генов, достаточно широко до этого распространенные в популяции. В этом случае вероятность встречи гаметы, несущей вновь мутировавший ген, с гаметой, несущей такой же ген, будет достаточно велика. Если мутация, дающая рыжую окраску шерсти, возникла в питомнике спаниелей, в котором многие животные уже несут один или два гена рыжей окраски, то каждый первый щенок, получивший мутировавший ген, может оказаться рыжим. Человек несет много летальных генов. Встретит ли новый рецессивный ген себе подобный ген, возникший раньше, будет зависеть от распространенности уже существующих генов этого типа и от частоты родственных браков в популяции, где возникла новая мутация.
Еще одно исключение представляют сцепленные с полом рецессивные мутации. Эти мутации, подобно гемофилии у человека, проявляются у каждого самца, получившего Х-хромосому с мутировавшим геном от своей матери. Таким образом, они обнаруживаются среди сыновей облученных самок или среди внуков, получивших мутацию через дочерей облученных самцов. Большое число сыновей облученных матерей или внуков облученных отцов погибают от сцепленных с полом рецессивных леталей.
Использовать сцепленные с полом летали для изучения мутаций у дрозофилы предложил Мёллер еще до открытия мутагенного действия рентгеновских лучей. С тех пор этот метод стал излюбленным методом исследования мутаций. Это объясняется несколькими причинами. Поскольку частота леталей значительно выше, чем частота видимых мутаций, их легче получить в количествах, необходимых для статистических сопоставлений. Кроме того, метод обнаружения сцепленных с полом леталей отнимает мало времени и достаточно эффективен. Дочери облученных самцов скрещиваются с отдельными самцами, и в их потомстве подсчитывают сыновей. У дочерей, получивших от отца сцепленную с полом деталь, будет в два раза меньше сыновей, чем у остальных дочерей. Разработаны особые остроумные приемы для того, чтобы избежать необходимости подсчета в таких семьях внуков-самцов.
В одной ставшей хорошо известной во всех лабораториях генетики линии мух самки, с которыми скрещивают облученных самцов, несут в одной из своих Х-хромосом сцепленную с полом деталь. Если облученный сперматозоид привносит вторую сцепленную с полом деталь, у такой самки совсем не будет сыновей. Это учитывать исключительно легко, так что можно без особого труда просмотреть большое число семей второго поколения. Поскольку для развития одного поколения дрозофилы от яйца до половой зрелости требуется приблизительно две недели, весь опыт, начиная от облучения самцов до просмотра их внуков, длится приблизительно месяц.
Помимо генных мутаций рентгеновские лучи вызывают разрывы хромосом. Когда хромосома разрывается, разорванные концы остаются в течение некоторого времени клейкими и может произойти одно из трех событий:
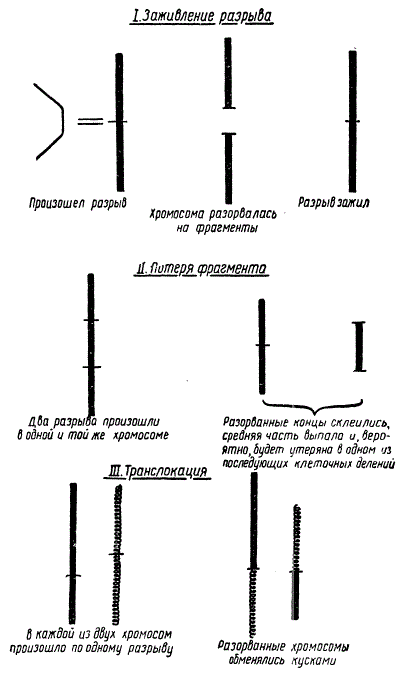
Разрывы хромосом под влиянием рентгеновских лучей и некоторые из их последствий
а) разрыв «заживает», т. е. разорванные концы могут снова соединиться по-старому;
б) разрыв «не заживает», и в ядре появляются обломки— фрагменты хромосом;
в) если в одной и той же клетке происходит несколько разрывов, разорванные концы могут склеиться по-новому, в результате чего образуются новые типы хромосом. Это называется хромосомными перестройками.
На рисунке показана «нехватка», т. е. выпадение и потеря куска хромосомы. Также видно транслокацию, т. е. обмен кусками между двумя разорванными хромосомами. Такой случайный обмен кусками не следует смешивать с обычным обменом одинаковыми кусками между хромосомами-партнерами, представляющим собой нормальное явление при делениях созревания. Транслокация может затронуть любые две хромосомы, независимо от того, являются ли они партнерами; обменные куски могут быть совершенно различной длины.
Посмотрим, как эти события, происходящие в облученных хромосомах, влияют на клетку и, если они происходят в гамете, на развивающийся зародыш?
а) «Заживший» хромосомный разрыв может вызвать генную мутацию, если один из генов, расположенных вблизи точки разрыва, поврежден этим процессом. Однако часто этого не происходит, и длительное повреждение не возникает.
б) Клетка, в которой облучение вызвало появление хромосомных фрагментов, ведет себя совершенно нормально до тех пор, пока она не начинает делиться. Однако хотя разорванная хромосома все еще способна образовывать свой двойник, механизм, при помощи которого новые и старые хромосомы распределяются по дочерним клеткам, выходит из строя в случае нарушения цельности хромосомы. Клетка, содержащая хромосомные фрагменты, часто погибает при попытке к делению. Даже если ей удается проделать одно или несколько делений, хромосомные фрагменты чаще всего теряются, в результате чего появляются клетки, не содержащие всех необходимых для выживания генов. Таким образом, незажившие хромосомные разрывы различными путями приводят к гибели делящейся клетки. В этом заключается одна из причин повышенной чувствительности к облучению тканей, в которых идет деление клеток, по сравнению с тканями, в которых деление прекратилось. Опухоли представляют собой быстро растущие ткани, содержащие много делящихся клеток; поэтому они гораздо легче повреждаются рентгеновскими лучами, чем окружающие здоровые ткани. Это явление широко используется в рентгенотерапии рака.
Если хромосомный фрагмент появился в яйцеклетке или сперматозоиде, оплодотворенное яйцо погибает обычно при одном из первых делений. Поскольку эта гибель происходит даже в тех случаях, когда только одна из гамет претерпела генетическое повреждение, его можно определить как доминантный эффект. Это явление называется доминантной летальностью в отличие от летальности, выявляющейся в последующих поколениях, которая вызывается рецессивными летальными генами. Опыты на животных показывают, что доминантная летальность представляет собой одно из важнейших последствий рентгеновского облучения. У дрозофилы из многих яиц не вылупливаются личинки в тех случаях, когда отец или мать подвергались облучению, несмотря на то что микроскопическое исследование подтверждает факт оплодотворения. Численность помета у мышей значительно уменьшается, когда самец или самка облучались. При высоких дозах рентгеновских лучей живые детеныши могут вовсе не родиться, несмотря на то что при вскрытии самки обнаруживали нормальное число оплодотворенных и приступивших к развитию яиц.
в) Смерть клетки и доминантная летальность могут возникнуть и в тех случаях, когда концы разорванных хромосом соединились неправильно, но это не всегда имеет место. Некоторые хромосомные перестройки не оказывают заметного влияния на особь, или по крайней мере этого влияния нельзя обнаружить без специальных генетических методов. Другие перестройки, как, например, потерю маленького кусочка хромосомы, нельзя отличить от мутаций, происходящих в генах, которые содержались в утерянном куске. Это объясняется тем, что многие мутировавшие гены утрачивают свою функцию, подобно гену альбинизма, который перестал стимулировать процесс образования пигмента. Вот почему часто невозможно определить, зависит ли нарушение развития от потери гена или потери функции все еще присутствующего гена.
Многие транслокации действуют как доминантные летали. Некоторые транслокации не затрагивают жизнеспособность своих носителей, но резко снижают их плодовитость, убивая половину внуков до того, как они вырастают. Детальное объяснение механизма, лежащего в основе этих явлений, завело бы нас слишком далеко. В общих чертах можно сказать, что транслоцированные хромосомы с трудом могут отыскать себе партнеров среди нормальных хромосом во время делений созревания, и это приводит к появлению гамет с неполным набором хромосом и генов. Транслокация и вызываемая ею уменьшенная плодовитость наследуются как доминантный ген. В результате облучения мышей было получено много линий, в которых половина потомков в каждом поколении всегда отличаются значительно пониженной плодовитостью.
Частота мутаций, вызванных рентгеновскими лучами
Дозы рентгеновских лучей измеряются в рентгенах (р), единица названа по имени открывшего их исследователя. Они измеряют число пар ионов, образующихся в данном объеме. Доза 200 р вызывает образование в два раза большего числа пар ионов в клеточном ядре, чем доза 100 р, а доза 400 р — в два раза большего, чем 200 р, и т. д.
Зависимость между дозой рентгеновских лучей и частотой вызываемых ими мутаций и хромосомных разрывов исключительно проста. Появление мутаций или хромосомных разрывов обусловлено ионизацией, возникающей в непосредственной близости от хромосомы или даже внутри нее. Вероятность этого возрастает пропорционально числу пар ионов, возникающих во всем ядре, точно так же как для плохого стрелка возможность попадания в цель повышается пропорционально числу выстрелов, которое ему разрешается сделать. Частота мутаций и хромосомных разрывов повышается и понижается, следовательно, пропорционально повышению или понижению дозы рентгеновских лучей. Так, например, когда самцов дрозофилы облучают в дозе 1000 р, в результате около 3% сперматозоидов несут сцепленную с полом леталь. При дозе 2000 р частота сцепленных с полом деталей возрастает до 6% и при дозе 3000 р — до 9%. Не следует забывать, однако, что эти частоты представляют собой средние величины и не в каждом опыте они получаются с абсолютной точностью, так же как в игральных костях шестерка не всегда выпадает на каждый шестой бросок, но на 600 бросков она выпадает приблизительно 100 раз.
Эго простое соотношение между дозой и эффектом справедливо даже при самых низких из испробованных доз (5 р), и нет основания сомневаться в том, что оно окажется правильным и для еще более низких доз. По-видимому, можно утверждать, что не существует столь низкой дозы, которая не обусловливала бы появления мутаций и хромосомных разрывов в количествах, пропорциональных числу образующихся пар ионов. Это очень важное обстоятельство, о котором следует помнить при обсуждении генетического влияния рентгеновских лучей на человека.
Для тех, кто интересуется последовательностью исследований, следует добавить, что для облегчения понимания вопроса при изложении нами зависимости между дозой и эффектом телега была поставлена впереди лошади. В действительности, пропорциональность, существующая между дозой и эффектом, и независимость эффекта от распределения ионизации во времени были установлены раньше, а на основании этих открытий был сделан вывод о зависимости появления мутаций и хромосомных разрывов от ионизации, возникшей по соседству с хромосомой.
Если любая ионизация, при условии ее возникновения достаточно близко от хромосомы, может вызвать мутацию или хромосомный разрыв, то определенная доза рентгеновских лучей будет вызывать их с одинаковой частотой независимо от того, получена ли она организмом в течение одного кратковременного и сильного воздействия, или в течение продолжительного и слабого воздействия, или же на протяжении нескольких воздействий. Это подтвердилось на примере мутаций, полученных в сперматозоидах дрозофилы.
Мёллер сравнивал число сцепленных с полом леталей, возникающих в сперматозоидах, получивших дозу 200 р при кратком интенсивном облучении за 8 минут и при очень продолжительном слабом облучении, продолжающемся более 30 дней; в этом последнем случае вместо рентгеновских лучей применяли γ-кванты радия. В обоих случаях было получено около 6% сцепленных с полом леталей. Тот же результат мог быть получен при облучении мух в течение одной минуты дозой 200 р на протяжении десяти последовательных дней. Таким образом, частота мутаций и хромосомных разрывов в сперматозоидах дрозофилы определяется исключительно величиной дозы, а не способом ее распределения во времени. То же самое справедливо для сперматозоидов мыши.
Сперматозоиды представляют собой особый тип клеток, в которых почти не идут процессы обмена веществ. По-видимому, в метаболически более активных клетках определенную долю повреждений, нанесенных хромосомами, можно исправить до того, как они реализуются в виде мутации или разрыва. Кроме того, эффективность этого восстановленного процесса зависит, вероятно, от интенсивности радиации, в силу чего исправление повреждений происходит в большей степени в условиях слабого хронического облучения, чем при коротких интенсивных воздействиях. В незрелых половых клетках самцов и самок мышей хроническое облучение у-квантами при мощности дозы 0,009 р в минуту вызывает в 4 раза меньше мутаций, чем такая же общая доза, примененная в виде острого воздействия при 90 р в минуту. Таким образом, для этих клеток одна и та же доза при хроническом облучении оказывается менее эффективной, чем при остром облучении. Однако даже при этом более низком уровне эффективности мы также обнаруживаем прямую зависимость между эффектом и дозой, когда каждое увеличение дозы в два раза приводит к удвоению частоты мутаций. По-видимому, не существует столь низкой дозы, которая не вызывала бы хоть какие-то генетические повреждения.
Нетрудно заметить, что при обсуждении отношения между дозой и эффектом были рассмотрены только генные мутации и хромосомные разрывы, а не такие перестройки, как транслокации, возникающие в результате склеивания разорванных хромосом. Это упущение имеет свои причины: закономерности соотношения между дозой, мутациями и хромосомными разрывами неприложимы к хромосомным перестройкам. Это вполне понятно. Хромосомные перестройки представляют собой вторичный эффект хромосомных разрывов, и для того, чтобы произошла перестройка, необходимо присутствие по крайней мере двух разрывов в одном и том же ядре. Совершенно справедливо, что число клеток с одним разрывом возрастает пропорционально числу актов ионизации в ядре, т. е. пропорционально дозе рентгеновских лучей.
Это, однако, не распространяется на ядра с двумя разрывами, частота которых возрастает быстрее с увеличением дозы и, напротив, становится непропорционально низкой при очень малых дозах.
Мы не будем рассматривать вопрос об установлении характера зависимости числа хромосомных перестроек, например транслокаций, от дозы рентгеновских лучей. Следует упомянуть только об одном обстоятельстве: поскольку в большинстве ядер концы разорванных хромосом остаются клейкими только в течение ограниченного периода времени, образование хромосомных перестроек в отличие от образования разрывов зависит от скорости, с которой следуют друг за другом отдельные события ионизации. Если промежуток между двумя ионизациями, вызывающими разрывы, слишком велик, первый разрыв окажется «зажившим» или потеряет свою клейкость до того, как произойдет второй. Даже большая доза рентгеновских лучей вызовет появление незначительного числа хромосомных перестроек, если она будет распределена на протяжении длительного промежутка времени.