Рассмотрим теперь такую группу явлений, когда наследование признаков связано с особенностями самой цитоплазмы, которые могут возникать в процессе развития организма под влиянием факторов внешней среды (онтогенетическая или фенотипическая предетерминация) под влиянием генотипа (генотипическая предетерминация).
Известен ряд фактов, когда наследование некоторых признаков материнской линии обусловливается изменениями в цитоплазме, вникающими в ней под влиянием определенных внешних агентов. Обычно такие изменения, появившиеся в одном поколении, являются нестойкими и через несколько поколений постепенно исчезают, возвращаясь к исходному типу.
А. Кюн в своей работе с наездником Habrobracon juglandis показал, что воздействие повышенной температурой на яйца самок до оплодотворения приводит к изменению окраски тела у их потомства. В последующих поколениях при размножении в нормальных температурных условиях это изменение постепенно затухает. Когда температурному воздействию подвергаются самцы, а самки выращиваются в нормальных условиях, подобного эффекта не наблюдается.
Сходные результаты еще ранее были получены В. Тоуэром в опытах по температурному воздействию на колорадского жука. Тоуэр высказал предположение, что в цитоплазме яйцеклеток еще до оплодотворения под влиянием повышенных температур происходят некоторые изменения, предопределяющие изменения в окраске тела потомства.
Кюн предложил для обозначения этого явления термин «предетерминация». В данном случае предетерминация обусловливается влиянием внешних агентов. Процесс затухания самого изменения признака в ряду поколений при возвращении организмов в исходные условия свидетельствует о том, что изменение было вызвано специфическим действием агента и не связано с репродуцирующимися элементами клетки. Подобные изменения относятся к так называемым длительным модификациям. Термин этот был предложен В. Иоллбсом в Г913 г., который провел в данной области обширные исследования на одноклеточных организмах.
В основе большей части случаев онтогенетической предетерминации, по-видимому, лежит приобретаемая под воздействием различных факторов внешней среды адаптация яйцеклеток, которая передается потомству по материнской линии при половом размножении и через соматические клетки в случае бесполого размножения. В некоторых случаях приобретенные в онтогенезе таким образом адаптации могут вызвать адекватные изменения признаков и создавать ложное впечатление наследования приобретенных признаков. Следует только иметь в виду, что такие адаптации варьируют в пределах нормы реагирования организма, обусловленной генотипом. Длительные модификации могут постоянно сохраняться в ряду поколений при условии сохранения вызывавших их факторов; при отсутствии последних происходит постепенный возврат к исходному состоянию.
Еще больший интерес представляют факты предетерминации свойств цитоплазмы под влиянием генотипа материнского организма.
Первые указания на наличие такой предетерминации были получены еще в прошлом веке в опытах Т. Бовери и других исследователей при изучении развития яиц различных беспозвоночный животных. В опытах по мерогонии и андрогенезу было установлено, что тип дробления яйца полностью соответствует тому типу, который характерен для материнского организма, несмотря на то, что материнское ядро отсутствует. Для объяснения этого явления было выдвинуто предположение, что в процессе образования яйцеклетки генотип материнского организма каким-то образом уже оказал влияние на формирование особенностей ее цитоплазмы, которые и обусловливают в дальнейшем характер дробления.
Ярким примером генотипической предетерминации цитоплазмы является наследование направлений некоторых пресноводных моллюсков, которые являются гермафродитами и могут размножаться как путем самооплодотворения, так и скрещивания. Встречаются два типа закручивания раковины: против часовой стрелки — левозакрученные (синестральное закручивание) и по ходу часовой стрелки — правозакрученные (декстральное закручивание). Направление закручивания раковины определяется одним геном; правозакоученность D доминирует над левозакрученностью d.
При реципрокных скрещиваниях гибриды F1, имеющие один и тот же генотип Dd по фенотипу различаются. В скрещивании DD x dd все гибридные особи имеют материнский тип — правозакрученные раковины. В скрещивании dd x DD, где у материнского организма раковина с левой закрученностью, потомство также имеет материнский тип завитка, т. е. левозакрученную раковину.
От самооплодотворения форм F1, гетерозиготных по этим генам (Dd) как в первом, так и во втором скрещивании, все потомки F2 обладают правозакрученной раковиной, хотя гибриды F1 (материнские формы) различались по фенотипу. Когда было исследовано потомство F3 от каждой особи в отдельности, то выяснилось, что 1/4 семей имели левый завиток, а 3/4 — правый. Простое менделеевское расщепление по данной паре признаков 3:1 выявилось, таким образом, не в F2, а только в F3.
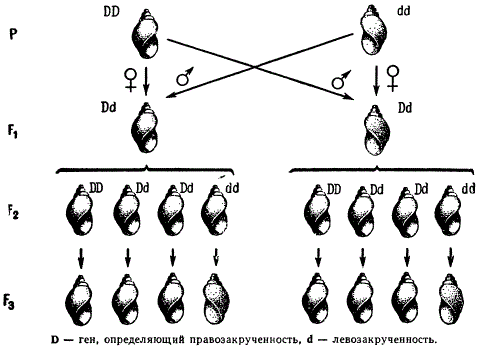
Роль цитоплазмы в наследовании направления завитка раковины у Limnaea
Такой тип наследования показывает, что фенотип потомков всегда соответствует генотипу исходного материнского организма (Р1), а не генотипам тех зигот F1, из которых они развиваются. Это может быть только при допущении, что данный признак предопределяется генотипом материнского организма в цитоплазме яйца в процессе его развития. Рассмотренный тип наследования и является в собственном смысле слова материнским. Под материнским типом наследования, называемым матроклинией, следует понимать предетерминацию цитоплазмы яйцеклетки генотипом материнского организма до ее оплодотворения.
Как было выяснено дальнейшими эмбриологическими исследованиями, направление завитка раковины определяется характером спирального дробления оплодотворенного яйца, т. е. расположением бластомеров по спирали вправо или влево, что в свою очередь зависит от ориентации веретена при втором делении дробления.
Описанное явление обнаружено у ряда животных; например, у шелковичного червя пигментация оболочки яйца определяется по тому же типу, что и характер завитка у улитки. У этого же объекта наследование вольтинности, т. е. смены поколений без диапаузы, также определяется особенностями цитоплазмы яйцеклетки, детерминированной материнским генотипом еще до оплодотворения.
Кроме данного типа предетерминации, имеет место также более длительное влияние генотипа материнского организма на развитие эмбриона с постепенным затуханием его в онтогенезе. Предполагается, что подобный тип материнского влияния широко распространен. Одним из ярких примеров такого влияния может служить наследование пигментации у мельничной огневки (Ephestia kuhniella).
У этой моли Кюн нашел мутантную форму, которая отличалась от дикой формы на стадии личинок и имаго. Рецессивная мутация в гомозиготном состоянии аа определяет красную окраску глаз у взрослой формы, неокрашенную гиподерму гусеницы и слабую пигментацию личиночных глазков; она изменяет также окраску головного ганглия и семенников у имаго. Дикий тип определяется доминантным геном А. У имаго этот ген обусловливает черную окраску глаз, коричневую окраску ганглия и семенников, а у гусениц — красноватую гиподерму и сильно пигментированные глазки. Было найдено, что в пигментации дикого типа принимает участие вещество типа «гормона» — кинуренин, предшественником которого в теле насекомого является триптофан.
Если кинуренин инъецировать личинкам мутантного генотипа аа, то они приобретут пигментацию дикого типа. Следовательно, у мутанта аа блокирован синтез кинуренина, а доминантный ген А контролирует участие кинуренина в синтезе пигмента.
Если скрещиваются особи, различающиеся по генам, АА х аа, то все их потомки F1 являются гетерозиготными Аа и все имеют окраску дикого типа. Анализирующее скрещивание АА х аа должно было бы дать расщепление в отношении 1Аа : 1аа, однако на стадии гусеницы особи обоих генотипов оказываются пигментированными и различаются лишь на стадии имаго. В реципрокном анализирующем скрещивании, где самка имеет генотип аа, расщепление и по фенотипу соответствует отношению 1:1.
Причина различия в проявлении фенотипа при данных скрещиваниях заключается в следующем. В теле материнского онанизма, имеющего генотип Аа, ген А продуцирует кинуренин, который частично попадает и в яйцеклетку. У особей аа запаса этого вещества хватает лишь на то, чтобы синтезировать пигмент на стадии гусеницы. Но по мере расходования кинуренина образование пигмента тормозится, так как особи генотипа аа не способны самостоятельно синтезировать кинуренин. Поэтому взрослые формы аа, появившиеся от скрещивания Аа х аа, не имеют характерной для дикого типа пигментации глаз, покрова нервных ганглиев и семенников.
Рассмотренный пример материнской наследственности не относится непосредственно к цитоплазматической наследственности, поскольку здесь свойства цитоплазмы детерминированы действием ядерных генов, а не элементами цитоплазмы. В примере с определением направления завитка у улитки первые фазы дробления яйца детерминируются геном матери. В опытах с мельничной огневкой гены материнского организма определяют синтез вещества, которое вкладывается в яйцеклетке и в последующем влияет на развитие пигмента у гусениц. В обоих примерах действует механизм хромосомной наследственности, который изменяет цитоплазму яйцеклетки еще до оплодотворения.
Как мы уже говорили, особенно тщательные исследования генотипической предетерминации провел Н. Н. Соколов у реципрокных гибридов между видами дрозофилы — Drosophila virilis, D. littoralis. Гибриды от скрещивания D. littoralis X D. virilis развивались нормально, а при реципрокном скрещивании почти (до 100%) обнаружили сильное нарушение развития и разного рта аномалии в строении крыльев, глаз, ног и т. д.
Цитологический анализ гигантских хромосом в клетках слюнных желез гибрида от второго скрещивания показал, что у него наблюдаются различные нарушения в наборе хромосом, которые являются следствием частой патологии митозов в его соматических клетках. Атипическое деление клеток было обнаружено также на ранних стадиях развития гибридных зародышей, чем и объясняются, по-видимому, уродства и аномалии у имаго.
Так как нарушения митоза не наблюдались у гибридов от первого скрещивания, то причину нарушений нужно искать в особенностях цитоплазмы этих видов. Однако в F2 оба гибрида независимо от направления скрещивания в делящихся клетках имели небольшое количество атипических митозов и были в этом отношении одинаковыми. Оказалось, что видовые различия цитоплазмы, проявившиеся у гибридов F1, в F2 исчезли. Сходство обоих гибридов F2 автор объясняет тем, что они происходят из яйцеклеток, сформировавшихся в самках F1 которые имели одинаковый генотип.
Чтобы выяснить роль ядра в предетерминации свойств цитоплазмы яйцеклетки, автор провел дополнительную серию скрещиваний. Гибрид D. littoralis х D. virilis, нормально развивавшийся в F1 повторно скрещивался с отцовской формой. После полного замещения у этого гибрида хромосом материнского вида хромосомами отцовского, достигнутого уже к четвертому поколению возвратных скрещиваний, была получена новая форма, самки которой при скрещивании с самцами D. littoralis давали в потомстве столь же высокий процент атипических митозов, как и гибриды D. virilis х D. littoralis.
В результате беккроссов цитоплазма полностью изменилась в сторону свойств цитоплазмы D. virilis под влиянием хромосом этого вида. В том же случае, когда в этом гибриде замещаются не все хромосомы материнского вида, свойства цитоплазмы даже к пятому поколению возвратного скрещивания изменяются не полностью.
Применение цитогенетического метода в исследованиях Н. Н. Соколова впервые позволило четко проанализировать генотипическую предетерминацию свойств цитоплазмы. В них показано, что именно генотипическая предетерминация определяет несовместимость цитоплазмы одного вида с хромосомами другого.