Выше уже указывалось, что Маркс и Энгельс отмечали в качестве большой заслуги Дарвина произведенный им материалистический анализ явлений приспособлений или органической целесообразности. Дарвин предложил убедительное причинное объяснение ее — теорию естественного отбора форм, которые обладают наиболее полезными в данных условиях изменениями.
Рассмотрим этот вопрос подробнее.
По Дарвину, приспособленная, или адаптированная организация составляет основу характеристики любого биологического вида. Приспособление (адаптация) есть такая особенность морфо-физиологической организации живой формы, ее индивидуального развития, ее реакций и поведения, которые обеспечивают существование в конкретных внешних условиях. В этом смысле организация любой формы целесообразна. В ходе онтогенеза, на всех его стадиях и фазах, организм достигает видимого соответствия условиям жизненной обстановки. Этой проблеме Дарвин уделял огромное внимание, так как путем причинного объяснения органической целесообразности достигается причинное объяснение эволюционного процесса.
а) Факты, доказывающие, что адаптация есть реальное, действительно наблюдаемое явление органической природы. Ряд других фактов рассматривается дальше.
б) До работ Дарвина эти факты толковались, однако, как свидетельство «скрытого умысла, или цели, обнаруживающихся в процессе созидания организмов» (Тимирязев). Адаптированность организмов сводилась (в скрытой или явной форме) к действию нематериальных: факторов, или к проявлению мудрости творца. Констатируя этот факт, крупный ученый XIX века Дюбуа-Раймон указывал, что «давно уже привыкли рассматривать эту проблему, как не решаемую естественным путем».
Итак, до Дарвина органическая целесообразность рассматривалась как одна из загадок природы.
в) Дарвин существенно изменил эту точку зрения. В его теории вопрос ставится следующим образом.
Приспособления возникают из случайных неопределенных изменений. Они исторически развиваются в рядах поколений путем постоянного устранения отбором всех менее приспособленных. Органические формы преобразуются шаг за шагом. В каждом поколении возникают различные изменения, разные, биологически неравные, формы. Одни из них выживают и дают последующие поколения, другие гибнут. Этот процесс продолжается и в последующих поколениях. Таким образом, достигнутое приспособление оказывается не результатом свойства или способности организмов изменяться в соответствии с условиями среды, а результатом выживания «избранных» форм. Можно сказать, что путь к приспособительному совершенству, при всей относительности последнего, устлан останками павших поколений.
Очевидно, приспособление не есть некоторое абсолютное внутреннее свойство данного организма. Наоборот, всякое приспособление оказывается лишь явлением относительного и временного соответствия (конкретного признака или физиологической особенности) конкретным условиям среды.
Понятие «приспособление» и «признак» не обязательно совпадают. Не всякий признак есть приспособление. В новых условиях определенные признаки могут сохраниться, но в то же время они могут потерять значение приспособлений. Следовательно, приспособление относительно, временно, преходяще, оно, повторяем, есть не свойство, а лишь явление отношений конкретных признаков к конкретным условиям.
Из этих фактов вытекает дарвиновское понимание различий между изменением и приспособлением. Изменения могут иметь различное значение: они или полезны, или вредны, или индиферентны. Изменение становится приспособлением только тогда, когда оно полезно. Такова постановка проблемы приспособлений у Дарвина. Ввиду ее важности рассмотрим ее подробнее. Прежде всего, если бы организмы обладали особым свойством или способностью всегда изменяться приспособительно, то нельзя было бы констатировать у них элементов нецелесообразного, или дистелеологического состояния, организации или поведения. В действительности же подобные состояния организации и поведения имеют место. Рассмотрим некоторые примеры.
1. Дистелеологический характер морфологических структур. Естественно, что ожидать среди диких видов частых случаев дистелеологических состояний нельзя. Подобные формы подвергаются естественному истреблению (элиминации).
Тем не менее, дистелеологические морфологические состояния безусловно существуют. Прежде всего сюда относятся многочисленные уродства. Не останавливаясь на них подробнее, следует подчеркнуть, что они возникают как под влиянием внутренних (эндогенных) факторов, так и внешних (экзогенных). И в том, и в другом случае уродливые формы возникают и развиваются в ходе онтогенеза. При этом уродства могут возникать как во внутренних, так и в наружных органах и их системах, приобретая в ряде случаев смертельное (летальное) значение. Самый факт возникновения уродства показывает, что органическая целесообразность относительна и что в самой живой материи нет каких-либо нематериальных «сил», которые направляли бы развитие организма непременно в сторону целесообразной структуры. Единственной такой силой — притом вполне материальной — является естественный отбор, устраняющий всех уродов, которые в данных условиях не жизнеспособны. Однако, нужно иметь в виду, что самое понятие уродства относительно. Некоторые уродства сравнительно жизнеспособны, хотя и должны во многих случаях рассматриваться как дистелеологические состояния. Было, например, установлено, что в населении некоторых видов брюхоногих моллюсков, обладающих нормальной правозавитой раковиной, наблюдаются уклоняющиеся (аберрантные) формы с левозавитой раковиной. Гаузе и Смарагдова (1939) показали, что левозавитые раковины брюхоногого Fruticicola lantzi, при прочих равных условиях, значительно менее жизнеспособны. Авторы полагают, что левозавитые формы обладают дефектом в ассимиляционных процессах и наряду с этим процесс потребления запасов протекает у них физиологически менее экономно. Особенности организации некоторых форм легко приобретают дистелеологическое значение. Так, например, у насекомых наблюдаются дистелеологические состояния при рождении личинок и во время личиночной линьки, а равно в период превращения во взрослую форму (imago). Личинки окаймленного плавунца отрождаются с нежными покровами белого цвета. Было показано (Blunck, 1916), что в это время они очень уязвимы и заметны и многие из них становятся жертвами рыб, лягушек, тритонов, и крупных насекомых. Когда личинки начинают линять, то не всякая линька проходит удачно. Многим особям линька вообще не удается. Нередки случаи, когда личинка не может высвободить ног или головы из старой кожи. Многие из них после линьки бывают настолько утомлены, что не могут достигнуть поверхности воды. Такие личинки поголовно задыхаются.
Своеобразные явления наблюдаются у некоторых низших рачков, например у Bosmina, Podon, Evadne и других форм. Попав на поверхность воды, они не могут вновь погрузиться в нее вследствие несмачиваемости кутикулы. Такое же явление наблюдается у некоторых свободно живущих пресноводных круглых червей и т. п.
Все же в естественной обстановке дистеологические состояния являются исключением. Совершенно другие отношения наблюдаются в тех случаях, когда организмы освобождаются из-под контроля естественного отбора. В этих случаях дистелеологическая организация наблюдается весьма часто. Таковы, например, отношения у одомашненных форм.
У мазаевских баранов наблюдается переразвитие, приводящее к ряду дистелеологических признаков. Так, спираль рогов закручивается столь круто вниз и внутрь, что затылок животного сильно сдавливается. Если у переразвитых баранов не отпиливаются концы рогов, то животные погибают примерно на 2—3-м году жизни. Мошонка мазаевских баранов настолько атонична, что свешивается далеко вниз и мешает ходить. Кожа у них легко повреждается. У переразвитого шортгорнского скота наблюдается понижение продуктивности, плохая молочность. Роды проходят крайне тяжело вследствие переразвития мускулатуры бедра. У некоторых английских свиней наблюдается укорочение черепа, приводящее к неравномерному развитию челюстей. Зубы укороченной верхней челюсти не совпадают с одноименными зубами нижней челюсти и животное плохо пережевывает пищу. Далее наблюдается сужение носовых каналов, чем затрудняется дыхание, животные плохо едят, кашляют и т. п. Дистелеологические явления известны также у собак. У французских бульдогов часто наблюдается закручивание рудиментированного хвоста на бок. Садясь на хвост, собака постоянно раздражает его ткани. Эта травма приводит к образованию кисты. У той же породы наблюдается склонность к злокачественным опухолям. У хохлатых падуанских кур теменные кости черепа вздуты и продырявлены, что всегда грозит повреждением головного мозга. Хохол у этой породы, намокая во время дождя, свешивается вниз, закрывая глаза. Хохол вообще сужает их зрительное поле. Л. Райт указывал, что некоторые формы хохлатых кур плохо переносят климатические условия открытых и влажных мест. В подобных местах «они мрут, как мухи». Хохол у цыплят голландских кур препятствует им своевременно замечать приближающуюся опасность, и они часто становятся добычей ворон и дневных хищных птиц. У некоторых пород голубей, например, у короткоклювых турманов, птенчики не могут пробить скорлупу яйца, а самки, с их маленькими клювами, не могут кормить своих птенцов. По указанию Дарвина, у польских голубей вокруг глаз сильно развита голая кожа. Иногда она так разрастается, что «один голубь… едва ли мог видеть зерна, подбирая их с земли». У замечательных японских иокагамских петухов хвостовые перья достигают необычайной длины: до 3,5—4 м. Такие петухи сидят в особых клетках с верхним светом, а хвост свешивается вниз и всячески охраняется. Когда петуха выпускают на прогулку, присматривающий за ним работник поддерживает хвост на весу, или же хвост наматывают на специальную катушку. У вуалехвостов и телескопов хвостовой и парные плавники в значительной степени утратили свою двигательную функцию, а тело — способность к быстрому скольжению в воде. При попытках быстро плыть вуалехвосты часто зарываются носом вниз, описывая как бы «мертвую петлю» и прекращают поступательное движение.
В условиях искусственного отбора описанные формы продолжают существовать. В естественных условиях они не встречаются, так как были бы немедленно уничтожены действием естественного отбора. Однако, самый факт появления подобных нецелесообразных форм показывает, что организм, сам по себе, вовсе не обязательно целесообразен. Когда органическая форма выключается из действия естественного отбора, у нее сплошь и рядом появляются признаки и особенности, не возможные в естественной обстановке. Наблюдения над культурными линиями плодовых мух (дрозофил) показывают, что они сплошь и рядом не выживают в естественных условиях. Из этих фактов видно, что органическая целесообразность не есть сущность живых форм, или их «основное свойство». Целесообразная организация создавалась в ходе отбора, через элиминацию нецелесообразных форм, в том числе и тех типов, которые могут возникать и существовать под защитой человека. Именно по этим причинам нецелесообразные признаки, возможные у одомашненных форм, элиминируются у диких видов.
2. Дистелеологическое поведение животных также далеко не редкое явление. Рассмотрим некоторые примеры. Почвенная нематода Rhabditis coarctata способна инцистироваться, причем, как правило, цисты прикрепляются к конечностям и даже к ротовым органам жуков. Так как жуки подвижны, то такое использование их полезно для нематоды, обеспечивая ее распространение. Было, однако, показано, что цисты прикрепляются иногда к голове личинок мух. Такие цисты не могут обеспечить распространения нематоды, так как личинки мух не уходят с места кормежки. Совершенно очевидно, что прикрепление цист к личинкам мух нецелесообразно.
Гусеницы прецессионного шелкопряда передвигаются скоплениями, причем каждая задняя гусеница идет по паутинному следу соответствующей передней. Фабр проделал следующий опыт. Некоторое количество гусениц было посажено на край стеклянного сосуда. Первая гусеница двинулась по круговому пути, имея перед собой последнюю, замыкавшую плотное кольцо гусениц. Двигаясь друг за другом, гусеницы описывали круг по краю сосуда в течение семи дней. И лишь вследствие случайного отклонения одной из них, непрерывное круговое движение было нарушено. Стабильность инстинктивных действий в описанном случае оказалась нецелесообразной, так как, следуя друг за другом, гусеницы были обречены на голод.
Куколки некоторых бабочек-стеклянниц (Sesiidae) сидят в ходах древесины. С приближением срока отрождения бабочек, они продвигаются к выходу, высовываясь до половины наружу. Перемещение обеспечивается движениями брюшка, снабженного поясами острых шипиков, и имеет приспособительное значение, облегчая выход бабочек, которые не могли бы выбраться наружу из длинного хода. В случае сотрясения дерева, куколки втягиваются обратно в свои ходы. Отрождение бабочек происходит тогда в глубине, они не могут выбраться наружу и поголовно погибают. Таким образом, реакция куколок, вообще говоря полезная, оказывается» весьма рискованной в период рождения бабочек.
Интересный пример дистелеологическсго поведения можно подчеркнуть из наблюдений Беклемишева (1930) над питанием личинок комара Anopheles. Беклемишев указывает, что личинки Anopheles поглощают пищу не избирательно, механически набивая свой кишечник взвешанными в воде частицами. Состав сестона (взвешанная в воде смесь органических и неорганических частиц), как указывает Беклемишев, «очень часто… может оказаться вредным или опасным для личинок Anopheles, делая непригодными для них те или иные водоемы». Таким образом, механическое поглощение любого сестона показывает, что выживаемость личинок определяется их отношением к среде, а не какой-либо внутренней «изначальной» целесообразностью.
Общеизвестный представитель полезных насекомых, яйцеед трихограмма (Trichogramma evanescens, откладывает яйцо в яйце озимой совки.

Трихограмма (Tr. evanescens)
Вместо гусеницы из яйца бабочки выходят молодые трихограммы. Этим пользуются для борьбы с совкой, выпуская трихограмм на площадях, пораженных вредителями. Было замечено, что при очень обильном выпуске трихограмм поражаемость яиц вредителя не увеличивается. Оказалось, что трихограмма откладывает свои яйца в яйца совок, уже зараженные другими трихограммами. Было выяснено, что трихограммы не отличают здоровых яиц от зараженных. Перезараженность яиц совок ведет к тому, что часть развивающихся в них трихограмм гибнет, среди остальных рождаются дефективные, например, карликового роста, с пониженной плодовитостью или вообще нежизнеспособные. Таким образом, в биологии самой трихограммы имеются дистелеологические черты, ограничивающие ее размножение.
Известно, что амфибии не переносят длительного пребывания на сухой, нагретой солнцем, почве. Тем не менее автор наблюдал, например, лягушек, которые пытались преодолеть железнодорожную насыпь в жаркий летний день. Многие из них. остались на насыпи и высохли под лучами солнца. Был описан случай, когда жаркий июльский день на накаленный черноземный грунт (температура на поверхности земли достигала 62°) вылезла зеленая жаба Bufo viridis и тут же высохла (Огнев и Воробьев, 1924).
Русская выхухоль Desmana moschata приспособлена к водному существованию и делает в берегах водоема норы, выходное отверстие которых открывается только в воду. В нормальных условиях это выгодно, так как обеспечивается полная безопасность от возможных наземных врагов и непосредственный контакт с водной средой и кормовой базой. Файст (1921) описал последствия зимних паводков, когда холодная вода заливает лед озера и берега, затопляя норы выхухоли. Строение нор становится причиной гибели животных, которые не могут выйти на поверхность земли, а уходят в воду, т. е. под лед, где задыхаются и гибнут. Следовательно, то, что было полезным в одних условиях, оказалось гибельным в других.
Описанные случаи также говорят против представления об органической целесообразности, как «основном свойстве» живых форм.
3. Неполнота или приближенность приспособительной организации. Если все же в подавляющем большинстве случаев организмы всесторонне приспособлены к своей жизненной обстановке, то эти приспособления обычно носят лишь приближенный, а не абсолютно совершенный характер. Приспособление, говоря словами Дарвина, «не может быть признано наилучшим из всех возможных при любых возможных условиях».
В очень большом числе случаев морфологическая организация живых форм настолько мало специализирована или так неспецифична, что по ней трудно определить свойственный им образ жизни. Показаны три вида свободно живущих нематод.
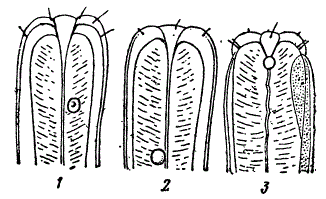
Три вида нематод из рода Monhystera. 1 — почвенная форма, 2 — пресноводная, 3 — морская (1,2 — по де-Ману, 3 — по Парамонову)
Одна из них живет в почве, другая в пресной воде, третья в полусоленых водах лимана, пограничных с морем. Тем не менее, их морфологическая организация совершенно однотипна. Подобные случаи не составляют редкости. Высокая специализация не достигается, если в ней нет биологической необходимости или если в данной среде нет более совершенных конкурентов. Например, у оляпки, как говорит Дарвин, «самый проницательный наблюдатель, исследующий мертвую птицу, не заподозрил бы подводных привычек, а между тем эта птица… существует только благодаря своей способности нырять, пользуясь своими крыльями под водой и цепляясь ногами за камни». Как известно, оляпка охотится под водой за личинками водных насекомых, моллюсками и ракообразными. Ее морфологические приспособления к подводным экскурсиям не специфичны, но, очевидно, иметь такую организацию биологически достаточно.
В ряде случаев морфологическая организация приводит даже исследователя к ошибочным выводам об образе жизни соответственных организмов. Например, подушкообразная форма растения не всегда свидетельствует об его приспособленности к холоду, так как эта же форма свойственна некоторым сухолюбам жарких областей. У животных сплошь и рядом обнаруживаются те же отношения. Американский крот Scalopus aquaticus имеет большие плавательные перепонки на лапах, откуда происходит и его видовое название. Однако последнее было дано Линнеем, не знавшим, что этот крот ведет не водяной, а обычный для кротов подземный образ жизни. Вообще об адаптивном значении признака можно судить только в том случае, когда выяснено его отношение к соответствующей среде.
Дарвин приводит в «Происхождении видов» целый ряд примеров. «Тот, кто верит, — писал он, — что все существа созданы такими, какими мы их теперь видим, должен порою испытывать изумление при виде животного, привычки которого не соответствуют его строению». Дарвин упоминает о горных гусях, которые обладают перепончатыми лапами, но лишь изредка приближаются к воде. Фрегат, как правило, не опускается к поверхности океана, «а между тем все четыре пальца у этой птицы соединены перепонками». Напротив, лысухи — типичные водяные птицы, а между тем «пальцы их только оторочены перепонкой». Водяная курочка хорошо плавает, хотя ее морфологический облик ничего не говорит об этом. Один из дятлов — Colaptes campestris имеет все типичные приспособления для лазанья по деревьям. Тем не менее, — пишет Дарвин, — «в некоторых обширных областях эта птица не лазит по деревьям и строит свои гнезда в норах по берегам рек», тогда как в других местностях этот же дятел «посещает деревья и выдалбливает дупло для своего гнезда в их стволах». Можно было бы привести очень большое число аналогичных примеров. Они показывают, что совершенство приспособлений в ряде случаев весьма приближенно, неполно, далеко от идеала. Приспособления никогда не бывают абсолютно совершенными, но зато, как в описанных выше примерах, они часто едва выражены и не специфичны. Эти факты также говорят против изначальной целесообразности, как «основного свойства», изначальной способности организма к целесообразным реакциям. Очевидно, описанные выше формы далеки от совершенства, хотя и осуществляют свои жизненные задачи. Весьма вероятно, что несоответствие их организации образу жизни свидетельствует о начинающихся или недавно возникших изменениях в этом последнем. При этом морфологическая организация еще че изменилась. Вообще в природе нет механически однотипного соответствия между организацией живой формы и средой ее обитания и ее образом жизни. Сплошь и рядом одни и те же жизненные задачи выполняются различными органами. Так, у некоторых растений крылья семян образованы за счет выростов прицветных чешуй (хвойник, Ephedra), у других — завязи (ревень и джузгун Calligonum), у третьих — в результате разрастания долей околоцветника и т. п.
Аналогичные факты широко распространены. Мы не можем создать трафаретной схемы адаптаций, которые имели бы универсальное применение. Очень многие высокогорные растения, например, приспособляясь к низким температурам, приобретают типичное строение холодолюбов: приземистый рост, распластанные и даже прижатые к земле (она не так быстро охлаждается) стебли и т. п. На Алтае, в зоне подвижных осыпей некоторые высокогорные растения скрывают свои длинные побеги под слоем угловатых камней. У холодолюбов наблюдается сильное опушение листьев и лепестков (эдельвайс). Многие растения приобретают форму плотной подушки, состоящей из скученных, связанных побегов. Одно алтайское растение Saussiurea involucrata обладает бледно зелеными листьями, свернутыми, как кочан капусты. Если их раздвинуть, то в пространстве между ними обнаруживаются корзинки темнофиолетовых цветов! Некоторые высокогорные растения не имеют всех описанных выше приспособлений. Их стебли вздымаются вверх, листья совершенно гладкие, общий морфологический облик не сходен с таковым холодолюбов. Тем не менее, их адаптации против холода весьма совершенны. Их ткани изобилуют обширными межклетниками, в которых сохраняется нагретый днем воздух.
Таким образом, приспособления к холоду оказываются неожиданно многообразными, осуществляясь самыми различными средствами.
То же и у животных. Как известно, хвостовое оперение птиц, имеет существенное значение в полете. У плохих летунов, в особенности у многих нырковых птиц, хвостовое оперение коротко и слабо развито. Однако в некоторых случаях слабое развитие хвостового оперения сочетается с относительно быстрым полетом. В этих случаях рулевую функцию хвоста берут на себя снабженные плавательными перепонками лапы (например, у чистика, Cephus grillea).
Следовательно, только экологическое наблюдение может точно выяснить функцию органа, и эта последняя в ряде случаев может оказаться совершенно неожиданной. Эти факты хорошо подчеркивают относительность приспособлений, отсутствие абсолютной, раз навсегда данной, шаблонной равнозначности между органом и его функцией в определенной среде.
Как правило, приспособление достигает лишь той степени совершенства, которая достаточна в данных условиях. Высокое совершенство приспособлений дельфина к водному образу жизни определяется совершенством плавательных приспособлений рыб, за которыми дельфин должен охотиться в условиях широкого морского простора. Он не может уступать им в адаптированности к водному образу жизни, а наоборот, должен превосходить их, иначе его питание окажется ограниченным.
Когда условия жизни не предъявляют столь высоких требований, приспособления не достигают и столь высокого совершенства, так как жизненные задачи выполняются и без них. Отсюда понятно и существование форм, организация которых неспецифична или даже, по видимости, не соответствует данной жизненной обстановке.
Это обстоятельство ясно указывает, что приспособление не дано заранее, как некое свойство организма, а определяется в каждом конкретном случае соответствующими условиями среды. Если, например, в данных условиях лов рыбы может быть достигнут без специальных приспособлений, то не будет стимулов и к их возникновению и развитию. Так, наша пресноводная кутора нападает на окуней, обладая едва выраженными приспособлениями к плаванию.
4. Приспособления, как выражение отношений между организмом и средой. Из сказанного выше видно, что мерой совершенства каждого приспособления является внешняя среда. Как правило, большинство приспособлений остается таковым лишь в совершенных конкретных условиях. Им свойственна пространственная и временная относительность. То, что является приспособлением в одних условиях, перестает быть им в других.
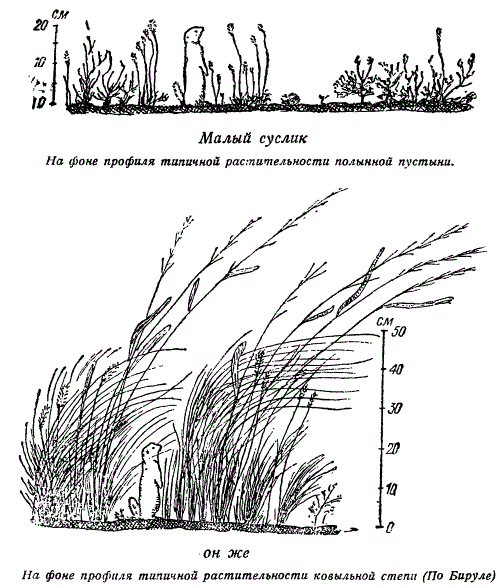
Исследования биологии малого суслика показали, что в условиях полынной полупустыни поведение этого грызуна построено по принципу: «первому заметить врага и своевременно уйти от него в нору» (Бируля, 1941). Это поведение в условиях, полынной пустыни целесообразно и отвечает интересам индивидуальной и коллективной защиты колонии сусликов от врагов. Тревожный свист одного из них служит предостерегающим сигналом для других. Однако исторически сложившееся поведение теряет свое адаптивное значение в густом и высоком травостое ковыльных степей, где труднее вовремя заметить врага и где сигнализация не оберегает и всю колонию. В новых условиях целесообразное поведение потеряло свою адаптивную значимость.
Приведем еще один наглядный пример. Личинки малярийного комара Anopheles maculipennis очень многочисленны в водоемах, заросших роголистником (Ceratophyllum) урутью (Myriaphyllum) и трехдольной ряской (Lemna trisulca), тогда как обилие плавающих рясок, а также кувшинок и белых лилий для личинок неблагоприятно. Если плавающие ряски покрывают, как часто наблюдается, все водное зеркало водоема, то затрудняется доступ атмосферного кислорода к личинкам. Было установлено, что если количество кислорода в воде ниже 4 миллиграммов на литр, то развитие личинок становится не возможным (Беклемишев, 1932). Часто наблюдается, что водоемы покрываются и заполняются различной водной растительностью. Следовательно, количественное распределение в них личинок неизбежно окажется очень пестрым, а степень их адаптированности в разных условиях весьма различной.
Физиологическая приспособленность к определенным условиям вообще играет огромную роль. Рисунок показывает пределы температурных лимитов жизни хлопкового долгоносика, его яиц и личинок. «Зона активности» жука замкнута в относительно узкие пределы и уже — 7° являются смертельной температурой для его яичек и личинок. Очевидно, что эти факторы лимитируют и распространение жука.
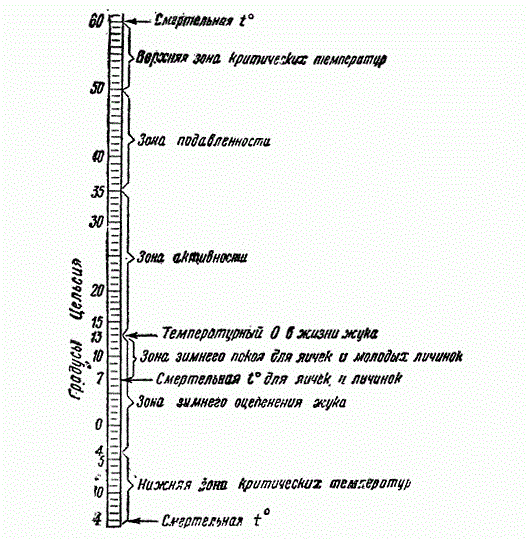
Шкала температурных лимитов жизни хлопкового долгоносика (Из Гёнтера)
Опаснейший вредитель сельского хозяйства, луговой мотылек характеризуется оптимумами и пределами тепла и влажности. Эти данные безусловно определяют в известной мере тот факт, что физиологическая приспособленность вида в разных частях ареала различна (по значению и степени). Следовательно, концентрация вида в разных частях ареала неизбежно будет пестрой, как и видно, например, на карточке распространения лугового мотылька. Таким образом, причины неравномерного распределения вида в границах его ареала определяются пределами его морфо-физиологических адаптаций. В смене этих связей большую роль играют также сами организмы.
Одним из частных проявлений влияния данного организма на адаптивную ценность признаков других организмов являются отношения между хищником и его жертвой. И в этих случаях наблюдаются изменения адаптивной значимости признаков. Например, столь действительное средство защиты, как колючий, иглистый покров ежа, не действителен против филина, который разрывает его на части и заглатывает куски тела своей жертвы вместе с иглами. Известно также, что сумчатый волк овладевает ехиднами и поедает их вместе с кожей и иглами. Следовательно, защитные приспособления этих животных теряют свое адаптивное значение при наличии указанных хищников, оставаясь действительными в отношении других.
Было показано (Беляев, 1927), что если окраска богомолов соответствует окраске растений, на которых сидят эти насекомые, то последние оказываются хорошо замаскированными. В этих случаях некоторые птицы — в опытах Беляева чеканы-каменки и пустельги — поедают преимущественно тех богомолов, окраска которых отлична от фона растений. Если же окраска соответствует последнему, то птицы истребляют богомолов лишь частично. Таким образом, совпадение окраски богомола с окраской субстрата, т. е. гомохромия, полезно и имеет защитное, адаптивное значение, несовпадение, или гетерохромия, — вредно. Однако адаптивное значение гомохромии было действительно, когда истребителями были чеканы и пустельга. Когда же на площадку налетели вороны, то они съели и гомохромных и гетерохромных богомолов, т. е. «приспособленных» и «неприспособленных». Адаптация не как свойство организма, а как выражение отношения, выступает здесь весьма явственно.
Относительность адаптаций была также хорошо показана Сюиннертоном (1919). По Сюиннертону, несъедобность или съедобность некоторых насекомых, выделяющих пахучие защитные экстракты, определяется не только свойствами последних, но также физиологическим состоянием птицы. Каждое насекомое защищено своей «несъедобностью» только в отношении тех птиц, которые в данный момент недостаточно голодны, чтобы есть его. При большем же голоде поедается и такое «несъедобное» насекомое. При меньшем — оно не берется и будет съедено более съедобное. Относительность адаптации здесь вполне явственна.
В описанных выше примерах была показана относительность приспособлений к условиям, изменяющимся в пространстве. Однако, уместно различать также относительность приспособлений во времени. Эта последняя доказывается фактами вымирания многочисленных видов, родов, отрядов, а в некоторых случаях — классов и даже типов. Некоторые данные по этому поводу уже приводились. Общей причиной вымирания видов является утрата приспособляемости к новым условиям среды, хотя, конечно, детали этого процесса в каждом отдельном случае — различны.
Выводы. Итак, фактический материал подтверждает дарвиновскую интерпретацию органической целесообразности. Эта последняя прежде всего, несомненно, существует. Приспособленность органических форм к их жизненной обстановке — бесспорный факт. Однако приспособленная или «целесообразная» организация не есть «изначальное свойство» организма. Целесообразная организация создается в ходе истории органической формы путем постоянного уничтожения (элиминации) менее целесообразных. Приспособления всегда относительны. В естественной обстановке постоянно наблюдается колебание адаптивной значимости признаков и физиологических свойств и даже переход приспособлений в свою противоположность. Правда, подобные случаи еще плохо изучены, и только поэтому их сравнительно мало обнаружено. Как говорит Дарвин, «согласно теории естественного отбора скорее представляется удивительным то, что не открыто еще большего числа подобных случаев отсутствия абсолютного совершенства».
Это замечание Дарвина показывает, насколько глубоко и методологически правильно понимал он явление органической целесообразности.