Основное значение в эволюции имеют мелкие наследственные изменения. Дарвин придавал им ведущее значение. Противники теории отбора указывали, что предположение об отборе мелких изменений не выдерживает критики, так как мелкие изменения бесполезны и не могут давать каких-либо преимуществ их обладателю. Кроме того, указывалось, что вообще эволюция не имеет приспособительного характера.
Если бы виды формировались на основе этого начала, то они различались бы по адаптивным признакам. В действительности виды часто различаются по неадаптивным систематическим признакам. Таким образом, получается, что вся теория отбора несостоятельна, так как: а) мелкие наследственные изменения не дают преимуществ и б) видообразование не идет по приспособительным признакам.
Рассмотрим эти вопросы. Шмальгаузеном (1940) было совершенно справедливо указано, что мелкие индивидуальные изменения могут свободно накапливаться в популяции вида именно ввиду их незначительности. В самом деле, мелкие мутации, раз возникнув, не вызывают резких нарушений установившейся корреляционной системы, в особенности, если они фенотипически проявляются на поздних стадиях эмбриогенеза, когда они не влияют на уже прошедшие фазы формообразования. С этой стороны проблема возникновения мелких мутаций и их накопления не содержит в себе чего-либо неясного. Возникает, однако, вопрос о том, могут ли мелкие мутации иметь полезное значение. Еще Тимирязев, а позднее Шмальгаузен (1940) напомнили, что естественный отбор идет не через выбор наилучших, а через элиминацию «неудовлетворительных» (Тимирязев). При этом элиминация последних происходит именно потому, что существуют и лучшие формы. Отбор идет, следовательно, по принципу «лучшее есть враг хорошего», т. е. протекает (в естественной обстановке) механически (по крайней мере у низших организмов). Именно на этом основании зиждется отбор полезных изменений. Однако, спрашивается: могут ли иметь преимущество мелкие изменения? Прежде всего следует остановиться на том, что собственно называется «мелким изменением». Так называются едва различимые морфо-физиологические изменения, улавливаемые нашими органами чувств. Многое здесь зависит от наблюдательности. Например, Мичурин и американский селекционер Бербанк различали такие мелкие изменения у растений, которые не улавливались другими наблюдателями. Таким образом, понятие о «мелком изменении» субъективно. Однако, его субъективизм выявляется еще в упущении того простого факта, что если данное изменение наблюдателям представляется мелким, то мы еще ничего не знаем о том, насколько оно мелко для самого обладателя его, т. е. растения или животного. Мелкое морфологическое изменение может оказаться связанным с весьма существенными физиологическими различиями.
Например, различия между расами северной сельди выражаются в мелких морфологических признаках, которые, сами по себе, не имеют адаптивного значения. При этом эти признаки так мелки, что различия между ними удается установить только методами вариационной статистики. Зато физиологические особенности различных рас сельди имеют явный адаптивный характер. Так, одна раса нерестится осенью при Т° = 10—13° и солёности — 3,0—3,5%. Другая, соответственно, в июле и августе, при солености, равной 3,5%, а третья требует Т° = 4—6° и солености воды 3,4%—3,3%. Повидимому, эти расы весьма чувствительны к определенной pH водной среды. Следовательно, их истинными расовыми признаками являются не мелкие морфологические признаки, а физиологические различия.
Таким образом, прежде всего нужно помнить, что морфологические различия между видами могут быть неадаптивными, но что их эволюционная диференцировка шла не по морфологическим признакам, а по приспособительным физиологическим свойствам (Кашкаров, 1939). Именно эти физиологические приспособительные различия и составляют сплошь и рядом содержание адаптивных различий между видами, хотя субъективно они и кажутся нам «мелкими» и неадаптивными в их морфологическом выражении. Таким образом с этой стороны виды, безусловно, различаются по адаптивным особенностям.
Посмотрим теперь, не могут ли и мелкие морфологические различия иметь адаптивное значение.
Дубовский (1941) исследовал адаптивное значение признаков различных видов ракушковых рачков (Ostracoda). Изученные виды различаются по мелким признакам, ко принадлежат к разным стациям — зарослям водорослей, песку и ракушечнику, наконец, к илам. Различия между видами касались: степени развития первой пары антенн, формы и количества щетинок на них, положения антенн (они могут быть загнуты на спинную или брюшную сторону), степени развития коготков второй пары антенн и некоторых других мелких признаков. Было выяснено, что все эти признаки связаны с образом жизни, т. е. имеют адаптивное значение, и что, следовательно, в данном случае, диференцировка видов происходила по адаптивным различиям.
Наибольший интерес представляет поставленный Дубовским эксперимент. Через длинный наклонный лоток пропускалась вода, которая стекала в подставленную в конце лотка сеть. В верхнюю часть водного потока лотка пускалось некоторое количество рачков (Cypria curvifurcata). Сила потока воды была больше той, которой могли противостоять рачки. Поэтому они смывались вниз. Однако, при действии лотка в течение 45 мин. было смыто около 50% всех рачков. Смытые рачки собирались, а затем исследовались наравне с несмытыми. Были также установлены причины несмываемости части рачков, причем оказалось, что часть рачков зарывается в грунт, что и позволяет им противостоять смыву.
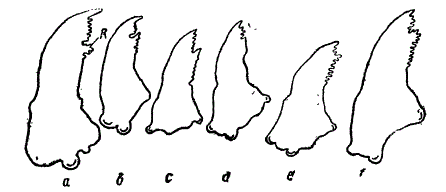
Правые мандибулы личинок из сем. Silphidae. а, b — хищники, c, d — падальники, e, f — всеядные, R — retinaculum.
Измерения показали, что «несмываемые» рачки обладают более короткими коготками первой антенны, короткими гребными конечными коготками первой ноги и т. п. У «смываемых» особей, которые не зарывались в грунт, указанные части несколько длиннее.
Различия настолько малы, что установить их, как достоверные, удалось только при помощи статистической обработки. Тем не менее, эти различия оказались, как видно из изложенного, адаптивными, причем было выяснено, что именно по этим мелким признакам Cypria curvifurcata распадается на два экотипа, которые распределяются в бассейне, в зависимости от силы (быстроты) водного потока. Адаптивная диференцировка в данном примере явственно совпадает с диференцировкой, имеющей эволюционное значение (т. е. на экотипы). Оба экотипа, как видим, различаются по мелким адаптивным морфологическим особенностям.
Подобные случаи, в конечном счете, повидимому, не столь редки. Нужны только, очевидно, не абстрактные рассуждения о значении мелких признаков, а проверка в опыте и наблюдении их адаптивного значения (Тимирязев).
Хорошим примером, рисующим адаптивное значение мелких различий, может служить также следующий факт. Жвалы (мандибулы) личинок жуков из семейства Silhidae различаются в очень мелких признаках — числом, расположением и размерами мелких зубчиков, которыми вооружена их вершина. Последняя диференцирована (Lengerken, 1938) на: а) «вершинный зуб», б) retinaculum и в) переднее озубленное пространство, отделенное от retinaculum глубокой вырезкой. На рисунке представлена правая мандибула серии видов Silphidae. Фигуры а, в изображают жвалы хищников. Retinaculum напоминает хищный зуб (dens lacerans) млекопитающих, а «вершинный зуб» — клык последних. У Silphidae из группы падальников «озубление» мандибулы дает переход от хищного типа к типичным «падальным» формам, с укороченным «вершинным зубом» (с, d). У всеядных Silphidae retinaculum округлена, «вершинный» зуб также округлен и в наиболее типичных случаях — укорочен (е, f). Эти мелкие различия, как видим, имеют большое адаптивное значение, так как связаны с образом жизни. Несколько более длинный «вершинный зуб» и несколько иное строение retinaculum имеют значение приспособительных различий, связанных с хищным, «падальным» или растительноядным образом жизни.
Из описанных примеров видно, что мелкие морфологические изменения, безусловно, могут иметь адаптивное значение.
Итак, мы пришли к следующим выводам:
1) Систематики различают виды по тем признакам, которые удобны для определения, независимо от того, являются ли эти признаки адаптивными, индиферентными или коррелятивными.
2) Признаки, используемые систематиками, часто не являются истинными видовыми признаками. Последними часто являются: а) тонкие физиологические различия, б) мелкие морфологические признаки малого систематического значения, но большой адаптивной значимости, в) в некоторых случаях «систематические» признаки могут совпадать с адаптивными (как выше у Silphidae).
3) Таким образом, истинные (а не только практически удобные) видовые признаки и свойства, несомненно, адаптивны. Эволюция видов идет по физиологическим и морфологическим различиям, часто весьма мелким по своему фенотипическому выражению, но весьма существенным по своему адаптивному значению. Эволюционное значение этих мелких изменений особенно велико потому, что они (как это видно в избранных выше примерах) не вызывают резких изменений в организации (сравни мандибулы хищных и травоядных Silphidae) и не нарушают общего плана ее, не разрушают установившихся корреляций и в то же время объективно приобретают большое жизненное значение.
Таким образом, представления Дарвина о ведущей роли мелких наследственных индивидуальных изменений находят свое подтверждение в фактах.