Первые детальные наблюдения, подтвердившие необходимость света для образования конидий и плодовых тел у темноцветных грибов, влияние света на их рост и пигментацию, принадлежат Leach (1962, 1963) и Trione et al. (1966), разработавшим методику выделения вещества Р310 — фоторецептора, ответственного за регуляцию образования конидиеносного аппарата у грибов Ascochyta pisi, Pyronema omphalodes, Pleospora herbarum, Alternaria chrysanthemii, Ophiobolus graminis. По предварительным данным, это вещество относится к пигментам флавиновой природы (Leach, 1965).
Позднее появилась серия работ, в которых испытано действие света на спороношение Botrytis cinerea (Tan, Epton, 1964a, b; Tan, 1975, 1978). При этом обнаружено угнетение образования спороношений под действием голубого света; лучи, близкие к ультрафиолетовым (400 нм), снимали этот эффект. Весь процесс спороношения впоследствии был разделен на шесть этапов. Облучение конидиеносцев на начальном или последнем этапе развития не угнетало образования конидий. Во всех остальных случаях действие голубого света вызывало образование уродливых стерильных конидиеносцев (Kumagai, 1978). Показана многократная обратимость этих процессов (Tan, 1975). Кроме того, освещение В. cinerea дальним красным светом снимало негативное действие голубого света. В свою очередь действие красного света снималось воздействием дальнего красного света. Таким образом, у гриба В. cinerea обнаружены фоторецепторы в длинноволновой УФ-области (340 нм) и 500 нм (голубой), а также при переменном освещении красным (600 нм) и длинноволновым красным светом (730 нм). Обнаруженные у грибов фоторецепторы называют микохромами. В результате проведенной работы (Tan, 1978) предложена однорецепторная модель микрохрома:
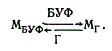
Кроме того, Tan предполагает возможное сопряжение двух микохромных систем:
где М* — предполагаемый микохром, БУФ — длинноволновые УФ-лучи (340 нм); Г — голубой свет 450 нм, К — красный свет 600 нм, ДК — длинноволновой красный свет 730 нм.
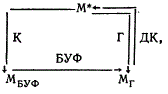
Первые шаги в определении природы микохромов предприняты Kumagai (1978). К настоящему времени микохромные системы, кроме В. sinerea, обнаружены у Alternaria tomato и Helminthosporium oryzae. Установлено, что 2-часовое облучение синим светом особенно губительно, если оно применяется после 6—8-часового выдерживания в темноте A. tomato и 4—8-часовой экспозиции в темноте В. cinerea. Новым этапом в изучении микохромных систем явились эксперименты на клеточных гомогенатах A. tomato. Центрифугирование клеточного экстракта в градиенте сахарозы позволило выделить три фракции. О действии УФ-лучей 305 нм (БУФ) на соответствующие фракции судили по изменению адсорбционных пиков на спектрофотометре. Из трех фракций максимум активности наблюдался в супернатанте, фракция 30 (в 30%-ной сахарозе) занимала промежуточное положение и наименьшей активностью обладала фракция 70 (в 68%-ной сахарозе). Максимальная активность после облучения синим светом наблюдалась в фракции 70, минимальная — в супернатанте. Таким образом, в микохромной системе удалось разделить два действующих начала — одно из них сосредоточено в основном в супернатанте и другое — во фракции 70. По гипотезе Kumagai, в грибе Alternaria tomato существуют два рецепторных пигмента, один из них (Пг) адсорбирует голубую часть спектра и является индуктором спороношения. Пигмент Пг накапливается в темноте, максимум аккумуляции у него наблюдается после 6—8-часового пребывания гриба в темноте. В результате адсорбции голубой части солнечного света, Пг переходит в Пбуф, способный адсорбировать свет 305 нм.
Схематически действие микохромной системы можно представить следующим образом:
![]()
Проверка этой гипотезы получена в экспериментах на клеточных гомогенатах. Из соответствующих фракций выделено два фоторецептора, причем обратимость адсорбции при освещении голубым и затем близкими УФ-лучами наблюдалась только если в реакционной смеси присутствовали фракция 70 и супернатант. Активное начало фракции 70 составляет пигмент Пг и супернатанта — Пбуф. Легкость, с которой Пбуф отделяется от Пг позволяет предположить, что он, по-видимому, растворен в цитоплазме и слабо связан с клеточными компонентами. Напротив, Пг по всей вероятности, тесно связан с цитоплазматической мембраной, так как не извлекается из клеток при обработке их ультразвуком или детергентами. Основные результаты были получены на изолированных протопластах A. tomato. Добавлением в реакционную смесь кислорода и дитионита удалось выявить, что пигмент обладал окислительно-воостановигельными свойствами. Характер действия Пг очень сходен с цитохромоксидазой, он отличается от нее тем, что восстановленная форма Пг не реагирует на добавление СО. Следует заметить, что только часть пигмента Пг реагирует на световое воздействие. Все изложенное позволило Kumagai (1978) дополнить первоначальную схему:
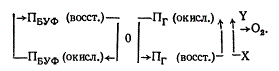
В приведенной схеме многое остается неизвестным — природа фактора, связывающего Пг и Пбуф, взаимодействие пигментов с молекулярным кислородом, прямое или посредством неизвестной реакции типа X — У.
Предстоит большая работа по выяснению механизма действия микохромной системы у грибов. Однако пока можно сказать, что 1) у грибов обнаружена регуляторная система, названная микохромной по аналогии с фитохромной системой у высших растений; 2) установлено участие микохромов в процессах образования конидиеносцев и конидий; 3) из клеточных гомогенатов Alternaria tomato, удалось выделить два фоторецептора, избирательно реагирующих на голубой (420 нм) и близкие УФ-лучи (305 нм); 4) более детальное изучение фоторецептора Пг позволило отнести его к пигментам. Пигмент тесно связан с клеточной мембраной и обладает способностью обратимо окисляться и восстанавливаться; в восстановленном состоянии он не реагирует на универсальный ингибитор дыхания — окись углерода. Действие микохромных фоторецепторов наиболее детально изучено на трех грибах: В. cinerea, A. tomato, Н. oryzae, два из которых относятся к меланинсодержащим.
О наличии микохромной системы предположительно можно судить по ингибированию спороношений при облучении синим светом и стимулированию — после облучения близкими УФ-лучами. Кроме указанных микохромных систем, установлено обратимое действие желто-красного и голубого света на спороношение гастеротмицета Sphaerobolus stellatus (Ingold, 1969; Ingold, Peach, 1970; цит. по: Tan, 1978).
Существует первичное и вторичное последействие света. К первичному относятся фотохимические реакции, которые происходят непосредственно после адсорбции света. Так, у Neurospora crassa флавиновый рецептор восстанавливает цитохром В; в микохромной системе адсорбент БУФ переносит энергию на Мг. Вторичное последействие света, по мнению Tan (1978), должно включать влияние света на метаболические процессы.
Освещение спорангиеносцев Phycomyces способствовало повышенному накоплению в них АТФ. Максимальное образование РНК и ДНК в конидиеносцах и конидиях N. crassa наблюдалось после освещения их, оно связано с циркадным ритмом гриба (Marteus, Sargent, цит. по Tan, 1978). Свет стимулировал процессы белкового синтеза, каротиногенеза у ряда грибов, активность окислительных ферментов (Tan, 1978). По данным М. К. Карабаева и В. Е. Петрова (1978), свет высокой интенсивности стимулировал усвоение СО2 пигментированными гетеротрофными микроорганизмами.
Характерным признаком фитохромной системы каротинсодержащего гриба Candida guillermondii является полная ее фотообратимость под влиянием света 730 нм (красного длинноволнового), если объект предварительно облучали красным светом (660 нм) (Фрайкин и др., 4973, 1974; Fraikin et al., 1976). Авторы получили результаты, свидетельствующие о наличии такой системы у каротинсодержащих дрожжей. Так, облучение светом 660 нм вызывало увеличение скорости деления дрожжевых клеток на 80—100%. Последующее освещение светом 730 нм полностью снимало это стимулирующее действие. Fraikin et al. (1976) установили, что продолжительность фотостимулирующего эффекта красного света не превышала 30 мин. Только в этот период времени фотостимулирующий эффект можно было снять освещением дрожжевых клеток С. guillermondii светом 730 нм. Кроме того, облучение предварительно освещенных светом 660 нм дрожжевых клеток сублетальной дозой УФ-лучей приводило к значительному увеличению их выживаемости. По мнению авторов, описанный процесс принципиально отличается от фотореактивации и фотозащиты, которые происходят при действии длинноволновых УФ-лучей и голубом освещении. Воздействие светом 730 нм не ингибирует эти процессы. Как правило,
фотореактивация и фотозащита продолжаются более 30 мин. Несколько позднее Valadon et al. (1979) наблюдали положительное влияние на процесс каротинообразоваиия кратковременного (10 мин) освещения красным светом гриба Verticillium agaricinum, которое полностью снималось последующим освещением светом 750 нм. Спектральная характеристика фракций пигмента, выделенных из мицелия гриба, оказалась сходной с таковой высших растений. На прорастание уредоспор Puccinia graminis положительно влияло 1-минутное освещение светом 660 нм; воздействие длинноволнового света 730 нм снимало этот эффект (Schneider, Murray, 1979).
Таким образом, углубленное изучение светового воздействия на грибы привело к открытию микохромных систем у представителей различных таксономических групп: аскомицетов, базидиомицетов, дейтеромицетов. Установлено регуляторное действие микохромов на скорость и характер спороношения (Botrytis cinerea, Alternaria tomato, Helminthosporium oryzae), процессы роста и деления клеток (Candida guillermondii), синтез каротиноидов (Verticillium agaricinum), скорость прорастания уредоспор Puccinia graminis. Высказаны предположения о механизме действия такой системы (Kumagai, 1978). Мы полностью разделяем мнение Schneider и Murray (1979), что меньшее количество фоторецепторов у грибов по сравнению с высшими растениями облегчит дальнейшее изучение их микохромных систем. Следует заметить, что подавляющее большинство грибов, у которых установлено присутствие микохромных систем, имеют пигменты каротиноидной и меланиновой природы.
Источник: Н. Н. Жданова, А. И. Василевская. Экстремальная экология грибов в природе и эксперименте. Киев: Наук. думка, 1982