Температурный режим почвы
Температура почвы зависит от количества солнечного тепла, поступающего на поверхность почвы и проникающего в ее глубину. На открытой поверхности почвы происходит наибольшее нагревание и наибольшая теплоотдача. Особенно велики максимумы температур на темноокрашенных и еще больше — на сухих песчаных почвах, где они подымаются до 60—70°, т. е. переходят через критическую точку существования живой плазмы (50—54°).
Минимумы на поверхности почвы более глубоки, чем в атмосфере, только при ясном небе и незащищенной почве, в ночное время или в ранние утренние часы. Но прохождение их в почву затормаживается, колебания суточных температур сглаживаются уже на глубине 25—50 см. Глубже 1 м проникают более длительные и интенсивные волны тепла и холода, вызываемые сменой сезонов года. Весной и особенно летом почва в среднем холоднее, чем воздух; осенью и зимой она теплее.
Важнейшие причины, определяющие особенности теплового режима, связаны с теплопроводностью и теплоемкостью почв. Влажные почвы более тепловодны и теплоемки, поэтому они (абсолютно или относительно) холодные. Сухие почвы и среди них особенно маловлагоемкие пески — теплые почвы.
Приводим средние температуры желтого сыпучего песка в 13 ч на поверхности и на глубине 10 и 25 см в течение летних месяцев (данные Олешских песков).
Средняя температура желтых песков в летние месяцы
| Глубина, см | Апрель | Май | Июнь | Август |
| 0 | 23,0 | 38 0 | 40 6 | 42,6 |
| 10 | 15,4 | 23,8 | 20,0 | 30,6 |
| 25 | 11,5 | 18,3 | 23,2 | 25,7 |
Таблица показывает, что во время наибольшей инсоляции (13 ч.) температура на глубине 10 см ниже, чем на поверхности почвы, на 8—15°, а на глубине 25 см — на 17—20°. Следовательно, верхняя часть корневой системы деревьев, особенно их горизонтальные корни, живут в более теплых, а глубинные корни — в более прохладных условиях. Кроны, стволы и корневые системы леса живут в разных фитоклиматических условиях; корни и стволы — в более мягком климате, кроны в более континентальном. Именно поэтому и возникла большая чувствительность корней как к высоким, так и низким температурам. Эту особенность корневых систем принимают во внимание при выкопке, перевозке и посадке деревьев на новые места. Корневые системы выкопанных сеянцев, саженцев и взрослых деревьев следует оберегать не только от действия солнечных лучей и ветра, но и от мороза.
Сомкнутый полог леса задерживает большую часть радиационного тепла. Именно на это явление прежде всего опирается давно установленное признание леса таким природным образованием, которое формирует свой особый лесной климат, свою особую лесную климатическую обстановку, отличную от климата соседней открытой местности.
Лесной полог отделяет лежащее ниже его воздушное пространство от свободной атмосферы и тем самым затрудняет обмен между ними воздухом и теплом. Внутри леса создается особый тепловой режим. Рассмотрим его, начиная с лесной опушки и стен леса.
Обращенные на юг стены леса и лесные опушки не дают тени. Отбрасывают наиболее длинную тень в течение почти всего светового дня южные опушки и стены леса, т. е. те, которые обращены на север. Поэтому южные опушки обладают наибольшим защитным действием на всходы и рост молодых сеянцев. Только в их тени в общих условиях избыточной инсоляции и недостатка влаги появляется густой самосев. Так, в сухих борах подрост сосны появляется на прогалинах, но только в тех их частях, на которые ложатся теки, отбрасываемые старыми соснами. Об этом явлении лесоводы образно говорят: «Подрост появляется в конусе полуденной тени». Здесь в ясные, летние дни температура верхнего слоя почвы градусов на 8—12 ниже, чем на открытых местах. Почва здесь более влажная, так как испарение в тени не так велико, как на открытом месте. В тени благоприятнее весь почвенный режим и менее опасны корнегрызущие вредители — личинки хрущей.
В более холодном и влажном климате и на влагоемких почвах оценка микроклимата южной (теневой) и северной (световой) сторон прогалины меняется. Так, северная сторона, обогреваемая солнцем с юга, значительно теплее южной, особенно в условиях мягкого морского климата. Это может стать ее преимуществом. На северной стороне легко поселяются и быстро растут светолюбивые породы — сосна, лиственница, береза и др. На южной стороне поселяются и хорошо растут теневыносливые породы — пихта, ель и другие. Но более теплолюбивым породам, в частности благородному каштану, дубу и буку, здесь может не хватать тепла.
Задержание тепловых лучей кронами препятствует нагреву лесной почвы и находящегося над ней слоя воздуха. Лесная почва на глубине 1—1,5 м в течение летнего времени холоднее, чем почва полей. Под сосновым лесом на глубине 60 см эта разница доходит до 2,7°, под еловым — до 3°, под буковым — до 3,2° (Ю. Шуберт, 1900). Зимой лесная почва, напротив, теплее, чем полевая, и не только под покровом вечнозеленых хвойных, но в еще большей степени под лиственными, накопляющими более мощный снежный покров. В среднем лесная почва холоднее на 1—1,5°, чем полевая, но в зимнее время она теплее на 0,5—1°. В лесостепной зоне европейской части Союза, за исключением Заволжья, почва зимой под дубравами не замерзает, так как защищена рыхлым снежным покровом и подстилкой из листьев дуба и его спутников. Только под ельниками, где из-за большого задержания снега кронами устанавливается тонкий снежный покров, почва промерзает на глубину до ½ м и больше. Но зато ельники сильнее умеряют высокие летние температуры, чем большинство лиственных пород.
Следовательно, почвенный климат леса мягче. Летом он прохладнее, зимой теплее, крайности температур сглаживаются. Незначительные различия средних температур не дают оснований для недооценки разницы между упоминаемыми явлениями, так как имеют большую длительность и многократную повторяемость.
Упомянутые особенности температурного режима лесных почв влияют на ход почвообразовательного процесса. Высокие температуры при недостатке влаги угнетают деятельность микрофлоры, замедляют разложение растительных остатков. Так, например, процесс выделения лесной подстилкой и почвой углекислоты (так называемое «дыхание почвы») в сухих борах почти полностью прекращается в жаркую и засушливую погоду, но возобновляется после каждого, даже и небольшого дождя. В противоположных условиях, на влажных и холодных почвах сырых боров, суборей и других типов леса, относящихся к торфянистым глееподзолам, разложение опада идет медленно. Подзолообразование и оглеение почв связано в лесах с временно избыточным увлажнением и недостатком тепла, способствующим задержке разложения растительного опада на промежуточных этапах. Избыток тепла при равномерном увлажнении почвы способствует ускоренной минерализации опада, Лесной полог, затеняющий почву и способствующий сохранению влаги в поверхностном горизонте, усиливает накопление кислого перегноя и подзолообразование в хвойных лесах; наоборот, в лиственных лесах подобный же тепловой режим благоприятствует накоплению мягкого (нейтрального) перегноя.
Осадки — первоисточник снабжения леса влагой. Твердые осадки холодного периода года — снег, иней, ожеледь, скопляясь на кронах деревьев, влияют на них своей тяжестью.
Осадки влияют на температурный режим леса. Летние осадки снижают температуру воздуха и почвы; зимние снежные осадки, наоборот, утепляют почву и занятый ими приземной слой воздуха.
Общее (годовое) количество осадков имеет значение для распространения лесов в крупных географических масштабах. Так, в лесных районах осадков бывает выше 400 мм, а безлесные местности — степи и пустыни получают в течение года менее 400 мм осадков. Связь между количеством осадков и распространением лесов детально изучил в конце прошлого века на североамериканском континенте лесовод Г. Майр. Он установил, что леса на этом обширном материке являются детищем влаги, приносимой воздушными течениями от Тихого и Атлантического океанов. Это происходит следующим путем. Насыщенные над океаном воздушные массы по мере продвижения внутрь материка расходуют свою влагу на образование осадков. Встречая на своем пути горные массивы и подымаясь вверх по их подветренным, обращенным к океану склонам, воздушные массы охлаждаются и отдают в виде осадков значительную часть своей влаги. Подветренные склоны в первой береговой зоне Тихого океана получают свыше 1000 мм влаги в год; в зоне Каскадных гор — 600—800 мм, в зоне Голубых и Скалистых гор — 400—600 мм. Переходя через вершины гор и опускаясь на их заветренные склоны, воздушные массы нагреваются; нагревание удаляет их от точки конденсации водяных паров, поэтому осадков на заветренных склонах выпадает мало. Следовательно, ближайшие к океану равнины и обращенные к нему склоны крупных горных цепей богаты осадками: центральные части континентов и обращенные к ним склоны больших горных массивов бедны осадками и в тем большей мере, чем они дальше расположены от океана. На первых развиваются леса, особенно роскошные — на океаническом побережье и ближайших к нему, обращенных к океану горных склонах. На вторых развивается травянистая и кустарниковая растительность прерий, степей, полупустынь и пустынь.
Г. Майр установил в качестве минимального количества для существования леса — 50 мм осадков в теплый период года, выпадающих в течение четырех летних месяцев, с мая по август.
Он обусловил этот показатель высокой относительной влажностью воздуха — около 70—75%. Однако в природе такая относительно высокая влажность редко сопровождает малое количество осадков. При условии, когда относительная влажность в летние месяцы падает до 50%, Г. Майр проводил границу леса по изогнете летних месяцев, равной 100 мм. При всех условиях указанные придержки являются грубо приблизительными.
Вопрос о существовании лесной омброхоры (границы леса, определяемой минимальным количеством осадков) детально проанализировал выдающийся советский лесовод-почвовед Г. Н. Высоцкий. Отдавая должное географическим исследованиям Майра, он отклонил в принципе существование лесной омброхоры, справедливо указывая на отсутствие во многих случаях непосредственных связей между количеством осадков и ростом леса. Так. во многих местах лесной (таежной), лесостепной и степной зоны количество годовых осадков колеблется от 350 до 500 мм, в пределах лесной зоны есть местности с 350 мм годовых осадков, а в пределах степной зоны — 450 мм и выше.
Г. Н. Высоцкий выдвинул понятие о лесной ксерохоре, т. е. о границе распространения леса, определяемой засушливостью климата. В качестве показателя влажности и засушливости климата Высоцкий предложил (1895) так называемый «омброэвапорометрический коррелятив» (ОК), представляющий собой отношение годового количества осадков к испаряемости по Вильду. Последняя величина определяется в миллиметрах влаги, испарившейся с открытой водной поверхности. Проводим несколько показателей ОК.

Осадки и испаряемость в различных географических пунктах, мм/год
ОК вдоль лесной ксерохоры близок к 1,0. Далее в пределах лесной зоны он становится больше единицы, а в пределах степной зоны уменьшается и доходит в среднеазиатских пустынях до 0,1. Показатель Г. Н. Высоцкого явился первой удачной цифровой характеристикой влажности климата, отражающей в одной цифре две ее главные функции — осадки и испарение. Последующие цифровые индексы Костяева, Ланга, Селянинова и др. лишь модифицировали принцип Высоцкого.
Однако нельзя удовлетворяться представлением о лесной ксерохоре как границе, проходящей по ОК=1,0. Как показал обширный опыт степного лесоразведения, искусственные леса из черешчатого дуба устойчивы и эффективны в зональных (плакорных) условиях всей степной зоны европейской части Союза, имеющей ОК от 1,0 до 0,5. Леса особого типа, состоящие из лоха, белой акации, гледичии и других засухоустойчивых древесных пород, а также довольно густые естественные «шибляки», «маквисы», «чанаррали», состоящие из засухоустойчивых видов дуба, сосны, фисташки, можжевельников и других пород, возможны и в полупустынной зоне Крыма, в среднеазиатских республиках, в Средиземье, в мексиканских пустынях и многих других засушливых областях, где соотношение осадков и испаряемости падает до 0,25. Другими словами, если не ограничивать представление о лесе тем его составом, который присущ южно-лесной подзоне и зоне лесостепи, если расширить это представление на низкопроизводительные леса и лесные заросли полупустынь и пустынь (в последнем случае они представлены зарослями саксаула), то древесная и лесная ксерохора отодвинется к ограниченным районам острозасушливых глинистых и каменистых пустынь.
Из всего изложенного следует, что при всех условиях степная зона Евразии, так же, как и области североамериканских степей и прерий, не могут быть отнесены к местностям, где лес отсутствует из-за сухости климата.
На земле в пределах суши нет максимальной лесной гигрохоры, т. е. географической границы, лимитирующей распространение леса из-за чрезмерного увлажнения. Граница леса с тундрой определяется недостатком тепла, который выступает ведущим в комплексе экологической среды; она является минимальной лесной термохорой.
Граница леса с пресными водоемами, в том числе и зарастающими, является эдафической и объясняется тем, что деревья. как и многие травянистые растения, не способны произрастать в водоемах из-за своей механической неустойчивости в этих условиях. Иными словами, они не растут на пресных водоемах из-за того, что не находят здесь стабильного твердого субстрата, на котором могли бы закрепиться. Именно поэтому на водоемы не идут ни выносливый к затоплению тополь, ни даже черная ольха, являющаяся болотным растением.
Лесные болота, покрытые низкорослой сосной или черной ольхой, широко распространены в равнинных областях, таких, например, как Припятское полесье. Равнинность рельефа уменьшает сток и способствует накоплению избытка воды. Количество осадков не играет при этом решающей роли. Гораздо большее значение имеет умеренная температура и малое испарение, а также наличие водоупорных горизонтов, которые способствуют высокому стоянию грунтовых вод.
В Сибири значительные пространства заболоченных лесов также связаны с равнинностью и в ряде районов — с неглубоким залеганием вечной мерзлоты в почве, в летнее время играющей роль водоупорного горизонта. Вечная мерзлота обусловливает в известной мере также и безлесие тундры, являясь одной из сторон общей причины ее безлесья — недостатка тепла.
Хотя на большей части нашей страны осадки вегетационного периода составляют свыше 50% от годовых, все же теплый весенне-летний период относят к периоду расходования влаги, в то время, как осенне-зимний называют влагонакопляющим. Это не случайно, так как низкие температуры осени и зимы способствуют сохранению влаги от физического и физиологического испарения, летом же на испарение расходуются как запасы предшествовавшего осенне-зимнего сезона, так и влага летних дождей.
Конец таяния снегов ранней весной является периодом наиболее глубокого промачивания почвы, наибольшего запаса почвенной влаги, возможного в течение всего года. В дубравах Лесостепи нередко бывают такие годы, когда после снежной зимы наступает засушливая весна и бедное дождями лето, однако прирост деревьев в этом случае не уступает приросту в годы, изобилующие летними осадками. Полезащитные лесные полосы в сухих степях могут удовлетворительно расти и не страдать от засухи, если они накапливают в течение зимнего времени некоторое количество снега, соответствующее по крайней мере минимальным потребностям деревьев во влаге. Аналогичные явления свойственны полеводству засушливой зоны. Снежные зимы обеспечивают большую влагозарядку и высокие урожаи сельскохозяйственных культур, имеющих короткую вегетацию.
Иную роль играет соотношение между зимними и летними осадками на песках. Так как пески маловлагоемки (полевая влагоемкость у них в 4—5 раз меньше, нежели у суглинистых почв) и водопроницаемы, то зимние осадки для лесов на глубоководных песках не имеют ценности.
Глубоководные пески отличаются от близководных в практике лесомелиораторов средним уровнем грунтовых вод 2,5 м.
Ранней весной при таянии снега большая часть влаги уходит в глубину песков за пределы корневых систем, если только она не была задержана близким к поверхности водоупорным горизонтом. Зато летние осадки, особенно от более значительных дождей (выше 5 мм), промачивают верхние слои песка и питают корешки. После продолжительного засушливого периода, что бывает чаще всего во второй половине лета, эффективными становятся на песках лишь дожди более 10 мм, так как для смачивания верхнего пересушенного слоя необходимо дополнительное количество влаги и некоторое гидростатическое давление, помогающее преодолеть несмачиваемость пересохшего песка. В противном случае вода уйдет на поверхностный сток или испарится.
Твердые осадки
Снег, изморозь, иней, ожеледь, град имеют немалое значение в жизни леса. Снежный покров в лесу предохраняет от вымерзания корневые системы, семена и всходы древесных пород, подрост, травянистые растения почвенного покрова, почвенную фауну. Рыхлый, пушистый снежный покров в лесу, будучи превосходным теплоизолятором, способен сохранять положительные температуры почвы в течение всей зимы. Мы уже отмечали, что почвы дубрав лесостепной зоны в зимы с рано ложащимся и устойчивым снежным покровом не замерзают. При этом лесная подстилка не играет большой роли, функцию теплоизоляции выполняет более чем на 90% снежный покров.
Многие теплолюбивые растения в климатических условиях умеренного пояса вымерзают. Таковы айлант, хурма, рододендроны, съедобный каштан и другие. Однако снежный покров сберегает им нижние части стволов и корни, в результате чего эти растения ежегодно могут восстанавливаться порослью от пня или корневыми отпрысками. Мороз как бы «стрижет» побеги теплолюбивых растений выше снежного покрова. В условиях постоянной морозной стрижки поколения некоторых растений способны существовать тысячелетия. Таково реликтовое растение — понтийская азалия, естественно распространенное в южной Белоруссии и северной Украине в качестве подлеска в суборевых сосняках-черничниках (В3 и В4). Лишь при наступлении значительных морозов до выпадения снега почва в лесу способна промерзнуть на значительную глубину и в таком состоянии при отсутствии оттепелей оставаться до весны. При этом теплолюбы могут значительно пострадать.
Снежный покров является ограничителем распространения лесной фауны. По А. Н. Формозову, северная граница ареала оленя проходит вдоль среднего годового максимума снежного покрова 40—50 см, так как снег большей мощности крайне затрудняет добычу корма. Для лося эта граница проходит при снежном покрове около 90 см. Дикий кабан отсутствует в северных частях лесной области из-за длительности снежного покрова и может быть еще в большей мере от промерзания почвы, затрудняющего добычу его привычного корма — корешков и корневищ травянистых растений. Снежный покров и ожеледь оказывают столь же отрицательное влияние на лесных птиц.
Зимнеголые лиственные породы, не подвергающиеся снеговалу и снеголому, страдают от ожеледи, т. е. от намерзания льда на поверхности ветвей. Особенно подвержены им густые дубовые жердняки, стволы которых и таких случаях поникают. От желеди и навала снега страдают тополевые насаждения всех возрастов; у тополей обламываются крупные ветки, а иногда и вершины. Наибольшее количество зависающего снега и намерзающего льда свойственно лесным опушкам, но опушечные деревья более приспособлены и менее страдают от этих явлений.
Потребление и транспирация влаги лесом для одной и той же породы зависит от бюджета влаги. Транспирация уменьшается при недостатке влаги в почве или, наоборот, увеличивается при ее большом количестве. Оптимальным количеством влаги является такое, которое покрывает полностью потребность растений и вместе с тем не создает в почве недостатка кислорода, необходимого корням для дыхания.
По отношению к увлажнению почвы, как и ко всякому другому экологическому фактору, следует, кроме оптимума, различать минимум и максимум, т. е. разные степени почвенного увлажнения, вызывающие качественно-разнообразный эффект в жизни леса. Особенно острым является минимум влаги — засуха. О ее наступлении свидетельствует увядание листьев, а в дальнейшем, при продолжающейся засухе, отмирание отдельных органов или даже целого растения от пересыхания. Различные древесные породы и их группы имеют разную степень устойчивости к засухе. Отличают ксерофитов (сухолюбов), приспособленных к большой потере влаги и подсыханию тканей, растущих в условиях недостаточного увлажнения; гигрофитов (влаголюбов), которые, напротив, очень чувствительны к засухе и растут лишь в условиях, обеспеченных влагой, выдерживающих недостаток кислорода в почве; мезофитов — средних по требовательности к увлажнению и по засухоустойчивости, растущих в промежуточных условиях увлажнения. Различают еще более дробные категории: ультраксерофиты — крайние сухолюбы, ультрагигрофиты — крайние влаголюбы (водяные растения) и переходные категории ксеромезофитов и гигромезофитов; первые стоят ближе к ксерофитам, вторые — к гигрофитам.
Кроме того, различают широту экологической амплитуды, которую проявляет растение, встречаясь не только в оптимальных для него условиях увлажнения, но и в других, соседних. Так, иногда ксерофитные растения, например, обыкновенная сосна, способны расти не только на засушливых («сухих») и оптимальных для нее «свежих» местообитаниях, но и на влажных, сырых и даже на торфяном болоте. При этом сосна лучше растет на почвах недостаточно увлажненных, чем на увлажненных избыточно. Ксерофиты с широкой амплитудой отношения к влаге носят название эвриксерофитов (эври — по-гречески широкий). Дуб, принадлежащий к ксеромезофитам, имеет также довольно широкую амплитуду отношения к влаге. Амплитуда дуба, правда, не столь широка, как у сосны, — дуб не идет на болота. Ксерофитами с узкой амплитудой являются саксаул, айлант, лох, пушистый и пробковый дубы и др.; гигрофитами с узкой амплитудой являются черная ольха и болотные ивы, мезофитами с узкой амплитудой — бук, явор, ильм, вяз, лещина.
Геоботаник Шимпер относил зимнеголые лиственные породы к тропофитам, т. е. к растениям, меняющим свой убор в зависимости от сезонов года. Во влажное время года, при выпадении значительных осадков и при наилучших условиях использования почвенной влаги, они покрываются листвой и ведут себя, как мезофиты или даже гигрофиты. В период зимней засухи, когда подача воды к листьям исключена, они теряют листья и становятся ксероморфными, т. е. приспособленными к отсутствию подачи воды из почвы, к сокращению испарения. Что касается вечнозеленых — хвойных, холодной и умеренной зон, то они также ксероморфны — их листья покрыты толстой кутикулой, имеют игольчатую форму, обеспечивающую им наименьшую поверхность, что является приспособлением для экономного испарения.
Русские ученые геоботаник Л. В. Гордягин и ботаник-физиолог Л. А. Иванов выдвинули в качестве важного показателя зимостойкости деревьев их зимнее испарение. Было показано в ряде опытов, что зимняя потеря влаги побегами древесных пород пропорциональна повреждаемости их морозами. К сожалению, и до сих пор мы не имеем достаточно ясной экологической оценки этих двух элементов зимостойкости деревьев: 1) прямых приспособлений к перенесению низких температур и 2) приспособлений к перенесению зимнего обезвоживания. Однако зимнее иссушение побегов играет хотя и не первостепенную, по в некоторых условиях заметную роль как в выработке зимостойкости, так и в явлениях суховершинности. Суховершинностью страдают чаще всего опушки леса, верхние части открытых склонов и другие ветроударные места.
Атмосферная засуха является результатом высокой температуры воздуха местного радиационного происхождения, отсутствия дождей, прихода сухих и нагретых воздушных масс со стороны. Почвенная засуха является результатом атмосферной засухи и сопровождающей ее сильной транспирации растений. При этом корни деревьев, поглощая влагу, исчерпывают ее запасы до степени прочно связанной, физиологически недоступной почвенной влаги, равной в песках 0,2—0,5%, в суглинках 5—10%. Эту последнюю долю почвенной влаги следует считать, как бы частью твердой почвенной фазы.
Длительные засухи могут быть катастрофическими для древесных растений. В наибольшей степени они сказываются в остро-засушливой и засушливой зоне юго-востока и юга нашей равнины. Здесь лесам в виде искусственных полезащитных полос присвоена важная функция «ветроломов» и снегораспределителей, способствующих смягчению засухи и поднятию устойчивости урожаев. Поэтому понятен интерес к вопросу о засухоустойчивости лесных насаждений в этой зоне.
Научные основы борьбы с засухой были заложены трудами выдающихся русских ученых — В. В. Докучаева, П. А. Костычева, К. А. Тимирязева, а в отношении леса в степях учениками Докучаева — Г. Н. Высоцким и Г. Ф. Морозовым.
Устойчивость против засухи — это способность растений переносить ее с наименьшим ущербом, в частности, с наименьшим снижением роста и урожая. Для полезащитных лесов засушливого юга главным является не столько прирост, сколько сохранение ими своей жизнеспособности и сопротивляемости по отношению к многочисленным вредителям из мира насекомых, охотно нападающим на подвялые листья и побеги, на ослабленные засухой деревья и кустарники.
Современное представление о ксероморфизме — признаках засухоустойчивости у растений является значительно более полным, чем прежнее, опиравшееся лишь на экономное расходование влаги у ксерофитов. Принято считать признаками ксероморфизма как у древесных, так и травянистых растений следующие особенности: толстые защитные (от испарений) листевые покровы, погруженные в них устьица, густое опушение листьев и побегов, редукция испаряющей части (мелкие листья или полное их отсутствие), восковой налет, способность регулировать движение замыкающих клеток устьиц, менять положение листьев по отношению к солнцу, сбрасывать листья при засухе, содержать в себе эфирные масла, также участвующие в регулировании испарений (С. П. Костычев), и др. Однако противопоставление способности интенсивно транспирировать по отношению к засухоустойчивости, представление об исключительной роли экономного расходования влаги для засухоустойчивости было отклонено работами отечественных ботаников — Н. А. Максимова, В. Г. Александрова, В. Р. Заленского. Б. А. Келлера, показавших, что очень многие ксерофиты и среди них засухоустойчивые древесные породы имеют даже большее по сравнению с мезофитами количество устьиц, более густую (в 3—5 раз) сеть листовых жилок и на этой основе большую интенсивность транспирации (иногда в 2—3 раза), чем у мезофитов.
Большое количество устьиц, хотя и более мелких по размеру, мощно развитая проводящая система необходима ксерофитам для энергичного газообмена, для продуктивного фотосинтеза, дыхания и транспирации, чтобы использовать короткий период вегетации в степях и пустынях, ограничивающийся ранней весной (пустыни) и простирающийся лишь на первую часть лета (степи). К ксерофитам упоминаемого типа относится большинство видов дуба, особенно жестколистные его формы — пушистый, пробковый, каменный и др.; степные кустарники — терн, шиповник, боярышник, береза, из тополей — наиболее засухоустойчивые среднеазиатские виды — тополь Болле и туранга.
В выработке засухоустойчивых приспособлений важную роль играет тип корневой системы. Так, на почвах малой влагоемкости, на песках и мелких каменистых почвах, особенно важным является выработка широкоразветвленной корневой системы, охватывающей значительный объем почвы. Выше уже отмечалось, что этому требованию отвечает сосна, развивающая длинные горизонтальные, а по ходу их часто отходящие вниз под прямым углом вертикальные корни, называемые якорными. Эти последние (а не стержневой корень) являются у взрослых сосен главными влагособирателями в засушливые периоды. Стержневые и косоуглубляющиеся корни преобладают у дуба, липы, лиственницы, ореха и других пород. Они повышают их влагоснабжение в засуху путем использования запасов влаги нижних горизонтов почвы и подпочвы, а в ряде случаев горизонтов грунтовых вод, расположенных хотя и на значительной, но досягаемой для них глубине.
Засухоустойчивость у древесных пород в степных и пустынных условиях повышается при более раннем начале вегетации, ибо весенний период в этих географических условиях обеспечен почвенной влагой. Так, в засушливых условиях смешанные посадки ранних и поздних дубов скоро превращаются в чистые из раннего дуба, вытесняющего своего конкурента по той причине, что к моменту распускания листьев поздней формы ранний дуб успевает использовать большую долю весенних запасов влаги в почве.
У древесных пород есть засухоустойчивые приспособления, позволяющие «обходить засуху». Такого рода приспособления неудачно называют «псевдоксерофитизмом». К ним относятся сбрасывание листьев в засушливый период, свойственное летнеголым лесам саванн. В наших условиях некоторые лиственные породы облегчают себе кризис летней засухи путем частичного, а иногда и полного сбрасывания листьев. К ним относятся тополи, особенно серебристый, сбрасывающий до ¾ своей листвы в июльскую засуху; мелколистный берест, белая и желтая акация (последняя может сбросить всю листву даже в конце июня), липа, гледичия, береза, лещина, ольха, ясенелистный клен и др. В качестве крайнего примера подобных приспособлений у нашей древесной флоры следует упомянуть тамариск, джузгун и саксаул, предельно редуцирующие свои листовые пластинки и с помощью этого радикального приспособления снижающие транспирацию в жаркий период.
Для борьбы с засухой в лесоводстве существует много разных, преимущественно агротехнических способов. Учитывая, что молодые одно-двухлетние сеянцы, особенно хвойных пород, крайне чувствительны к засухе, лесоводы сосредоточивают борьбу с ней на лесокультурной площади. Значительно повышает приживаемость и рост посадок в южных широтах подбор устойчивых к засухе пород — сосны, дуба, особенно — раннего дуба, успевающего использовать наибольшие в степной зоне весенние запасы почвенной влаги. В полезащитных лесных полосах южных степей устойчивыми и быстрорастущими являются мелколистный берест, белая акация, гледичия, американский клен, серебристый лох, абрикос и другие породы.
С особенной остротой почвенная засуха проявляется на песках. Важнейшими агротехническими приемами для увеличения и сохранения влаги в песчаной почве являются ее рыхление, глубинное внесение торфа, ежегодная 2—3-кратная культивация междурядий в течение по крайней мере первых 3—5 лет с тем, чтобы конкуренты за влагу — сорняки каждый раз возможно полнее удалялись.
Как показал обширный опыт, лучшие результаты посева и посадки лесных полезащитных полос в засушливых степях связаны с применением широких 3—5-метровых междурядий, постоянно культивируемых и используемых под пропашные культуры. Этот прием способствует накоплению влаги, созданию ее резерва для снабжения деревьев в засушливый период.
Накопление влаги в почве при создании лесных полос обеспечивается их собственным снегозадерживающим влиянием. Однако сильное снегонакопление в самих полосах невыгодно, поскольку в этом случае снег отнимается у защищаемых полей. Для устранения чрезмерного снегозадержания в полосах и равномерного распределения снега в межполосном пространстве применяют специальные рыхлые, ветропроницаемые структуры лесных полос. При этом наличие свободных (широких) влагонакопляющих междурядий становится особенно полезным для деревьев.
В ряде условий, где тракторная обработка неприменима (летучие слабозаросшие пески, склоны оврагов, скальные условия), лучшим способом посева или посадки леса является гнездовой. Его преимущества в этих условиях: лучшее использование неоднородностей почвы, быстрое создание первичных очагов лесной обстановки, сочетание выгодных сторон редких и густых посадок, в частности — расширенной площади питания при быстром смыкании крон внутри гнезд.
Избыток влаги в почве и заболачивание
Избыточное увлажнение отрицательно влияет на рост большинства древесных пород. Только черная ольха представляет собой настоящий гигрофит, ибо находит для себя самые лучшие условия на торфяных болотах низинного типа. Здесь уровень грунтовой воды располагается на 0,5—1 м от поверхности, выходя в весеннее время на самую поверхность почвы. Ольху сопровождают иногда (на самых богатых торфах) ясень, относящийся к особой болотной форме, подлесок из черной смородины, крушины и др. Однако эти породы не относятся к настоящим гигрофитам, подобно черной ольхе и ее травянистым спутникам — болотным растениям — калужнице, папоротнику, ирису и др. Корневые системы у них плоские, поверхностные, в то время как у ольхи и ее спутников — болотных растений — корни заходят в грунтовые воды и там разветвляются.
Особенностью гигрофитов является то, что они одновременно и ризоанаэробы, т. о. имеют такие приспособления в области корневой системы, которые дают им возможность выносить недостаток или даже полное отсутствие свободного (газообразного) кислорода в воде. Так, у черной ольхи сильно развиты чечевички на ветвях, молодой коре ствола и корнях, а также система анастомозирующих широких межклетников, проходящих из ствола в корни. По этой системе осуществляется воздушный обмен между атмосферой и живыми клетками молодых корешков, несущих функции поглощения влаги и минеральных питательных веществ. Кроме того, ольха использует азот воздуха с помощью микробов — азотофиксаторов, поселяющихся на ее корнях.
На болотах верхового типа растет сосна, иногда — с березой, но обе эти породы не гигрофиты: сосна — ксерофит, береза — мезофит. Поэтому деревья этих пород растут на верховых болотах невысоко и развивают только горизонтальную, плоскую корневую систему, углубляющуюся в торф всего на 20—30 см. Их преобладание объясняется недостатком минеральных веществ в верховом торфе, не позволяющем расти здесь черной ольхе, породе хотя и очень гигрофильной, но гораздо более требовательной к почве, чем сосна и береза.
На переходных к болотам влажных и сырых минеральных почвах, где сосна, ель, дуб, береза и другие породы достигают значительной высоты, а корневая система из-за близости грунтовых вод неглубока, взрослые насаждения часто страдают от ветровала. Во время бурь крупные деревья падают и своей тяжестью вырывают из почвы ту половину своей корневой системы, которая расположена со стороны, противоположной падению.
В естественных насаждениях, на почвах влажных и сырых, почти каждое дерево закапчивает свою жизнь ветровалом. Поэтому в таких лесах вырабатывается характерный микрорельеф корневых выворотов: ямы глубиною 20—40 см, шириной 1—3 м чередуются с бугорками той же высоты, но несколько меньшей ширины. Ямы образуются на месте вырванной корнями почвы, а бугорки — рядом, на месте ее осыпания, происходящего после сгнивания корней ветровального дерева. Впоследствии на понижениях разрастаются мхи, папоротники и травянистые растения — гигрофиты, а на возвышениях — мезофиты. Самосев древесных пород поселяется в виде групп на буграх, где он не страдает от вымокания. Вообще на почвах сырых и заболоченных самосев ели и других пород лучше растет на всяких повышениях, в том числе и на гниющих пнях, упавших стволах, на ветвях гниющих деревьев. Главная причина этого — лучшая аэрария субстрата, менее обводненного, чем почва.
Корневые системы большинства древесных пород, особенно хвойных, находят оптимальные условия водоснабжения и дыхания в условиях полевой влагоемкости почвы (НВ — по Роде). При этом увлажнении в пространстве между почвенными частицами, кроме воды, помещается достаточное количество воздуха, не менее 30% от общей порозности почвы. В горизонт капиллярного подъема грунтовых вод, где воздух остается в количествах меньших 30%, корни заходят лишь на небольшую глубину, ограниченную пределами сезонного колебания верхней границы капиллярной каймы. Сюда относится большинство лиственных пород, а из хвойных — ель, туя, болотный кипарис и др. Сосновые корни не выносят даже и этих условий; хотя они и врастают в верхнюю часть капиллярного горизонта, но ежегодно отмирают в нем.
Черная ольха заселяет корнями не только весь капиллярный горизонт, но и верхнюю часть горизонта грунтовых вод. Ее корни способны оставаться живыми в обстановке круглогодичного затопления, т. е. при состоянии полной влагоемкости почвы. Однако это возможно лишь в том случае, когда надземная часть ольхи — ствол и ветви не подвергаются долговременному затоплению. Затопление надземной части, имеющее место во время весеннего половодья в поймах рек, выдерживают древовидные ивы (главным образом, белая и ломкая) и тополя, особенно — черный и серебристый. Названные породы способны сохранять жизнеспособность и даже вегетировать при уровне полых вод до 5—6 м выше шейки корня в течение 2—3 месяцев, причем ивы образуют на затопленной части стволов дополнительные так называемые воздушные корни. Ольха, вяз и дуб выдерживают более короткие сроки затопления. Первые — до 1—1,5, третий — до 1 месяца. Береза, ель и сосна выдерживают затопление 2—3 недели. Совсем не выдерживают пойменных условий скальный дуб, граб и бук.
Деревья легче выдерживают затопление холодной водой. Теплая вода усиливает жизнедеятельность тканей, особенно дыхание. При недостатке кислорода дыхание ускоряет кризис, заканчивающийся отмиранием корневой системы от задыхания, гибелью всего дерева.
Для суждения о требованиях древесных пород к влаге были применены разные критерии. В середине прошлого века немецкие лесоводы Клаубрехт и Теодор Гартиг в качестве критерия требований к влаге использовали количество потери воды листьями в течение суток. Они получили шкалу, которая начинается с ольхи, теряющей влагу в наибольшем количестве, и заканчивается сосной и елью, стоявших на последнем месте по этому признаку. Подобная же шкала была получена Теодором Гартигом на основании весовой потери воды лиственных деревцев в водной культуре. Эбермайер составил шкалу по количеству золы в листьях, полагая, что зольность листьев тем выше, чем больше влаги они испарят. Он получил следующие ряды: ясень, ольха, клен, ильмовые, тополь — 7—10% золы; бук, дуб, граб — 4—5%; лиственница, пихта, ель — 2,9—3,5%; обыкновенная и черная сосна — 1—2%.
Долгое время наиболее авторитетными по данному вопросу считались данные австрийского ботаника Гёнеля, который определял транспирацию влаги у 5—6-летних деревцев, посадив их в сосуды с землей. Опыт сопровождался изоляцией поверхности почвы в сосудах от испарения, в силу чего потеря сосудов в весе могла быть приравнена к расходу влаги на транспирацию. Опыт длился в течение трех вегетационных периодов. Был получен следующий ряд древесных пород по убывающему количеству испаряемой ими воды на 100 г сухого вещества листьев (т. е. по интенсивности транспирации): ясень, береза, бук, граб, вяз, дуб, остролистый клеи, ель, сосна обыкновенная, пихта, сосна черная. Гёнель подтвердил вывод, полученный предшественниками, о большей потребности во влаге лиственных: по Гёнелю — сосны, например, испаряют в 6—8 раз меньше, чем дуб.
На основании данных Гёнеля в последующем вычисляли потребность и расход влаги разными насаждениями. При этом расход для дубовых насаждений в год на 1 га при 5 т сухого вещества листьев составляет 275 мм, что близко к данным, полученным Тростянецкой лесной опытной станцией, расположенной в лесостепной зоне. Г. Н. Высоцкий, придерживаясь данных Гёнеля, полагал, что сосняки испаряют всего около 40—50 мм в год на 1 га (1926). Данные А. А. Молчанова показали, что трапспирационный расход сосняков в 5—8 раз больше того показателя, который выдвинул Г. Н. Высоцкий, опираясь на данные Гёнеля.
Г. Н. Высоцкий в 1893 г. в Велико-Анадоле применил способ взвешивания живых листьев, оторванных от побегов в течение часа и получил следующие потери (%) содержания влаги в листьях и ее потери от первоначального веса листьев:
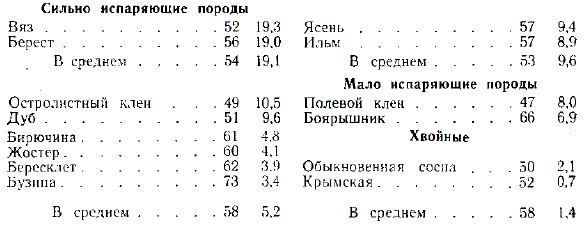
Г. Н. Высоцкий сделал вывод, что листья вяза и береста, вследствие тонкости их кутикулы и отсутствия значительного опушения, теряют влагу наиболее быстро, листья же ильма, благодаря значительному опушению, испаряют меньше. Кустарники и хвойные породы теряют мало влаги и не могут быть серьезными конкурентами дуба в степных посадках.
В позднейшее время советскими исследователями было получено много дополнительных данных, характеризующих потребность древесных пород во влаге. Кроме тою, лесоводы еще раньше, исходя из наблюдений над географическим распространением древесных пород на равнинах и в горах, отношением их к разным элементам рельефа, уровню грунтовых вод, поведению их при засухе или, наоборот, при переувлажнении почв, составили шкалы требования древесных пород к влаге. Они имеют существенное преимущество над описанными, так как составлены не по одному руководящему признаку, а по целому комплексу взаимно-контролирующих и дополняющих друг друга признаков.
Г. Ф. Морозов, опираясь на исследования Л. А. Иванова, подчеркнул, что обоснованное представление о потребности древесных пород в отношении влаги может дать лишь совокупность признаков и, в частности, совокупность понятий об интенсивности, продуктивности и экономности транспирации. Первая величина измеряется количеством воды на единицу площади (или веса) листьев, вторая — на количество всего органического прироста дерева, третья — на количество начального запаса влаги в листьях или во всем растении. Так, сосна и ель, близкие по интенсивности испарения, отличаются по его экономности — сосна в 2 раза экономнее ели. Сосна и дуб в 5-летнем возрасте испаряет 75% содержащейся в них воды, ель и бук 150—200%, т. е. две последние породы должны в течение суток дважды восполнять запас воды в своих органах.
Г. Ф. Морозов выдвинул понятия потребности и требовательности древесных пород к влаге. Потребность — это количество влаги, требуемое растением для удовлетворения его нужд, поддержания тургора его тканей, нормального хода физиологических процессов фотосинтеза, дыхания, защиты от перегрева, обмена веществ между разными органами растения и т. п. Требовательность — это отношение к условиям влажности почвы, уменье взять необходимое количество влаги в тех или иных почвенных условиях.
Потребность во влаге является одним из обязательных элементов, слагающих требовательность. Так, например, сильно потребляющая влагу черная ольха всегда растет на влажных почвах и не может расти на почвах сухих. Между тем интенсивно транспирирующие тополя и осина не нуждаются в чрезмерно увлажненных почвах и способны удовлетворительно расти на почвах, обеспеченных влагой лишь в течение весны, например на песчаных поймах, где после весеннего разлива уровень грунтовых вод падает на 2—3 м ниже поверхности почвы. Следовательно, черная ольха имеет не только большую потребность, но и более требовательна к влаге, чем тополя.
Г. Ф. Морозов указывал, что потребность во влаге сосны, ели и пихты невелика и примерно одинакова, но эти три породы сильно различаются между собой по требовательности к почве в вопросе обеспечения их влагой: сосна, обладающая очень широко разветвленной корневой системой, нетребовательна, в то время как ель и пихта с их слабее разветвленными, экстенсивными корневыми системами требовательны к влаге. Сосна является ксерофитом, а ель и пихта — мезофитами.
Для ели необходимо предоставлять почвы, влажные в самых верхних своих горизонтах, или поселять ее во влажном климате. Кроме того, ель и пихта имеют густое охвоение и большее количество хвои, которая держится на дереве у ели 5—8 лет, у пихты 8—12 лет, а у сосны — всего 2—3 года. Следовательно, потребность во влаге у ели и пихты больше в соответствии с испаряющей поверхностью кроны. Кроме того, под их густой полог проникает значительно меньше атмосферных осадков, а это также повышает их требовательность к влажности почвы и климата.
Лиственница и береза характеризуются высокой интенсивностью транспирации, гораздо большей, чем ель, бук и пихта. Однако малое количество хвои и листвы у первых и большое количество у последних меняет местами эти породы по количеству общего расхода влаги: густооблиственные теневыносливые породы оказываются более требовательны к ней, чем ажурно-кронные, светолюбивые. Следовательно, итоговая экологическая характеристика отношения растений к влаге может быть осуществлена лишь по их требовательности, а не только по одной потребности. Соотношение между листовой и корневой поверхностью, суммарная величина проводящих путей в древесине, размеры самого растения — все это измерители его требовательности к климатическому и почвенному увлажнению.
Приводим шкалу отношения древесных и кустарниковых пород к влаге, составленную на основе упомянутых признаков, а также на материале лесоводственного опыта. Породы в шкале и в пределах каждой группы расположены по нарастающим степеням требовательности к влаге.
Группа 0. Ультраксерофиты. Саксаул. Можжевельники. Фисташка. Миндальное дерево. Пушистый дуб. Пробковый дуб. Грабинник. Абрикос.
Группа 1. Ксерофиты. Крымская сосна. Обыкновенная сосна. Сосна Банкса. Айлант. Лох. Облепиха. Скумпия. Степные кустарники. Лохолистная груша. Абрикос. Мелколистный вяз. Самшит. Ива-шелюга. Белая акация. Японская софора.
Группа 2. Ксеро-мезофиты. Черешчатый и сидячецветный дубы. Груша. Берека. Черноклен. Остролистный и полевой клены. Берест. Гледичия. Черешня. Яблоня.
Группа 3. Мезофиты. Липа. Граб, Ясень. Орехи. Лиственница. Бук. Съедобный каштан. Конский каштан. Бородавчатая береза. Осина. Кедровая и веймутова сосна. Пихта. Дугласия. Ильм. Амурский бархат. Лещина. Бузина.
Группа 4. Мезо-гигрофиты. Вяз. Черемуха. Осокорь. Козья ива. Серебристая и ломкая ива. Пушистая береза. Ломкая крушина. Птерокария. Серая ольха.
Группа 5. Гигрофиты. Болотный экотип ясеня. Ива болотная серая, ушастая, лапландская. Кустарниковые березы. Айва. Черная ольха.
К шкале можно было сделать добавление шестой группы — ультрагигрофитов, к которой относятся в наших климатических условиях лишь водяные растения, не имеющие в своем составе древесных пород. В эту группу можно зачислить деревья мангровых лесов, растущих в субтропической и тропической зонах.