Эффект Варбурга — ингибирование выделения O2 и ассимиляции CO2 по мере увеличения концентрации O2 в среде — широко распространен у растений с циклом Кальвина.
Он обусловлен конкуренцией CO2 и O2 за субстрат ввиду проявления РБФК свойств, характерных для карбоксилаз и оксигеназ. Фотосинтез даже при нормальных концентрациях CO2 и O2 в среде приводит к образованию не только ФГК, но и гликолата.
Фотодыхание — окисление гликолата в глиоксилат на свету через редуктивный ПФП с усиленным потреблением кислорода и выделением CO2 — накладывает отпечаток на темновое дыхание. При этом излишки электронов фотосинтетической системы выводятся из цепи и используются для синтеза АТФ. Не вызывает сомнения наличие светозависимого потребления O2 у части растений как результат подавления темнового дыхания при фотосинтезе. Однако фотодыхание не всегда оказывается связано с фотосинтезом. Так, фотодыхание наблюдается у бесхлорофиллоносных мутаций хлореллы на сильном свету, которые способны синтезировать только каротиноиды. У таких мутантов фотодыхание составляет 50% от уровня темнового дыхания. В зеленых клетках при отключении фотосинтеза дыхательным ядом также происходит потребление O2 и выделение CO2.
При фотодыхании в 3—5 раз быстрее выделяется CO2, чем при темновом дыхании (М. Зелитч, 1976). Получается так, что, с одной стороны, в процессе фотосинтеза фиксируется CO2, с другой — она тут же теряется на 50% при фотодыхании. Природа здесь оказалась как бы не на уровне защиты интересов своего произведения. И среди растений С3-типа фотосинтеза обнаружены мутанты с блокированным фотодыханием.
В связи с подобными наблюдениями возникает вопрос, насколько С3-растения могут обходиться без фотодыхания. При осуществлении фотодыхания преобладает оксигеназная функция РБФК. При естественных концентрациях CO2 и O2 в атмосфере соотношение между фотосинтезом и фотодыханием у С3-растений составляет 3:1. Поскольку соотношение указанных газов в среде (в том числе и в клетке) часто меняется под влиянием температуры и времени суток (из-за колебания концентрации прежде всего O2), постольку необходимым оказывается переключение функции РБФК на оксигеназную функцию, и наоборот. Все зависит от того, какой газ находится в избытке.
Образование гликолата служит началом фотодыхания. Дальнейшее его превращение происходит в хлоропластах, пероксисомах и митохондриях. У большинства растений гликолат в хлоропластах образуется при падении концентрации CO2 и увеличении концентрации O2. Фотодыханию отводится некоторая положительная роль в обеспечении клеток промежуточными веществами (глутамат, γ-глутаровая кислота, глицин, серин, глицерат и т. п.), важном для дальнейших синтезов. Все это свидетельствует, что фотодыхание нельзя рассматривать как бесполезный или вредный процесс.
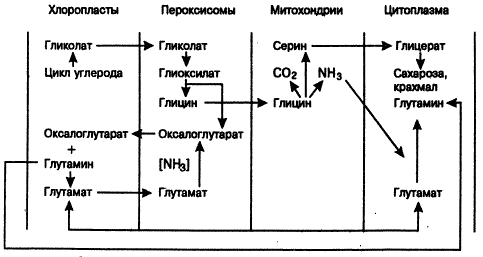
Фотодыхание способствует поддержанию постоянства CO2 внутри листа. У растений С4-пути существует другой механизм поддержания концентрации CO2 (фиксация его ЩУК). Фотодыхание тесно связано с общим метаболизмом в зеленой клетке, оно часто усиливается при уменьшении потребности растений в продуктах фотосинтеза и, в принципе, направлено на поддержание активности ферментативных систем и функции хлоропластов и митохондрий (А. Т. Мокроносов, 1983). Его роль иногда связывают с метаболизмом азотистых соединений, переносом их из одной органеллы в другую, а также с возможностью ограничения Накопления фосфогликолата.
Явление фотодыхания открыто в 1955 г. И. Деккером. Оно в большей мере выражено у растений С3-пути фотосинтеза и сопровождается потерей углерода в цикле Кальвина из-за снижения способности РБФК фиксировать CO2 и повышения ее активности окислять субстрат (РБФ) в присутствии O2 (А. Лайск, 1977). В случае окисления субстрата РБФК выступает в роли РБФ-оксигеназы. На примере рода Panicum, в пределах которого встречаются виды с С3-, С4— и промежуточными путями фотосинтеза, выявлены различия в содержании органелл. У P. riculare (С3-путь) клетки обкладки содержат в среднем 3 хлоропласта, 1,1 митохондрий и 0,3 пероксисом; у Р. milioides, Р. decipiens и Р. schenckii (промежуточные луга) органелл соответственно 10,6,17,7 и 3,2; у Р. priontis (С4-путь) в клетках обкладки органелл не только больше, но они и крупнее. У указанных видов фотодыхание снижается в направлении увеличения числа органелл в клетках обкладки (R. Н. Brow et al., 1983).
Начальные стадии фотодыхания (образование гликолевой кислоты) характерны для всех организмов с циклом Кальвина. Виды же различаются механизмами метаболизма гликолевой кислоты. Так, фотосинтезирующие бактерии и цианеи окисляют гликолат и синтезируют глицин. Цианеи содержат фермент гликолатдегидрогеназу и способны поглощать гликолат (в темноте и на свету), но утилизируют его слабо. У эукариотических водорослей в процессе фотосинтеза гликолевая кислота выделяется в окружающую среду, хотя у них несколько лучше выражен механизм утилизации гликолата (при участии фермента гликолатдегидрогеназы). Подобное выделение представляет собой способ защиты цикла Кребса от ингибирования и на начальных этапах развития жизни было необходимо для усиления симбиотических отношений бактерий и цианей с гетеротрофами (А. М. Степанова, А. А. Шумилова, 1980).
У высших же растений с С3-путем фиксации CO2 фотодыхание осуществляется наиболее сложно — путем полного превращения гликолевой кислоты, с образованием ряда важных промежуточных продуктов (аминокислот, порфиринов и др.). Дело в том, что с переходом растений к наземным условиям существования возможности выделения гликолата в среду ограничиваются ввиду невымываемости его из организма. Поэтому защита цикла Кребса от ингибирующего действия глиоксилата могла быть достигнута только путем полного превращения его в гликолат.
Несмотря на эти новшества, у высших растений потери углерода при фотодыхании составляют 50% от его ассимилированного количества. У них фотосинтез падает главным образом из-за конкуренции O2 и CO2 за молекулу РБФК и ее субстрат. Интенсивность фотодыхания у растений С3-типа возрастает при повышении концентрации O2 и температуры среды. Это ведет к ингибированию фотосинтеза в результате снижения сродства РБФК к CO2, особенно в аридных условиях. Поэтому отбор способствовал формированию растений С4-типа, у которых хорошо развита система концентрации CO2 (ФЕП, органические кислоты и др.) и выражена активность РБФК (при одновременном подавлении оксигеназной функции указанного фермента). Благодаря подобным и другим изменениям у растений С4-типа потери углерода при фотосинтезе невелики и гликолат накапливается, медленно.
Представляет интерес оценка роли фотодыхания в онтогенезе. Как известно, в экстремальных условиях водоснабжения устьица закрываются и резко возрастает фотодыхание. Это имеет значение для зашиты хлоропластов от разрушения (путем снабжения CO2) и сохранения нормального соотношения НАДФ/НАДФ ∙ Н2, в результате чего поддерживается процесс фотосинтеза и. растения предохраняются от истощения и теплового шока. В то же время образуемый H2O2 при взаимодействии O2 с ЭТЦ хлоропластов может способствовать отравлению тканей. Во избежание этого часть его также используется в хлоропластах для неэнзиматического окисления глиоксилевой кислоты. Возникающий на промежуточных стадиях фотодыхания АТФ метаболизируется в самих митохондриях, где его концентрация резко падает из-за ограничения оттока триозофосфата из, хлоропластов и поступления CO2 в ткани. Фотодыхание возрастает по мере старения растений и листьев (А. Т. Мокроносов, 1981).
Фотодыхание устраняет и Избыток первичной энергии, не используемой в фотосинтезе (J. Р. Gaudillera, 1982). Полагают, что само по себе фотодыхание не могло служить объектом отбора, а возникло в процессе соотбора как неизбежное следствию эволюции механизма фотосинтеза. Фотодыхание — адаптация к современному содержанию O2 и CO2 в среде. Содержание O2 в атмосфере (21%) превышает его потребность для темнового дыхания (2%). И фотодыхание предотвращает тормозящее влияние высокой концентрации O2 на физиологические процессы клетки и поддерживает целостность органелл путем дополнительной его утилизации для фиксации CO2 и окисления гликолевой кислоты. Итак, наблюдаем ряд событий, ведущих к усилению фотодыхания — выделение гликолата и развитие механизма его использования, появление микротелец (пероксисомы), усиление взаимодействия клеточных компартментов для обеспечения распределения и потока метаболитов. Эти события имели значение для обеспечения энергетического, баланса клетки, образования ряда промежуточных продуктов метаболизма, поддержания осмотического и протонного потенциала путем синтеза органических кислот, защиты клетки от токсического действия O2 и хлоропластов от повреждения «выбросом» CO2 из фотосинтетического цикла, устранения излишков фитогормонов в клетке путем их окисления, окисления жирных кислот и т. д. Значение каждого из указанных приобретений для выживания растений не вызывает сомнения.
Ведущее место в фотодыхании принадлежит пероксисомам. Ни один другой органоид клетки не выполняет столь разнообразные функции, как микротельца (А. У. Игамбердиев, 1988, 1990), в том числе и при генерации новых путей биохимических превращений. Фотодыхание, хотя и рассматривается как ответвление фотосинтетического пути, играет важную роль в обеспечении нормального функционирования Цикла Кальвина, снабжая его CO2 в обход реакций карбоксилирования. Оно также устраняет ингибирование ЦТК метаболизацией гликолата. По этим и другим причинам необходимо было выработать механизм утилизации гликолата, от которого ранее (водоросли) избавлялись выделением в среду. С появлением пероксисом у части водорослей (хлорелла) окисление гликолата в глиоксилат с образованием H2O2 имело значение для рассеивания свободной энергии, поглощенной клеткой. Выделение гликолата в среду его вымыванием имело значение для развития симбиотических отношений автотрофов и гетеротрофов, но оно не могло получить дальнейшего развития в наземных условиях, где исключалась возможность этого вымывания. Отсюда и необходимость отбора растений в направлении усиления утилизации гликолата в энергетических целях. Косвенно эти механизмы способствовали усилению транспорта воды, минеральных и органических веществ у сосудистых растений благодаря формированию вакуолярной системы. Интересно отметить, что в случае повторного перехода цветковых к обитанию в воде они начинают выделять гликолат в среду.
По-видимому, роль фотодыхания наиболее велика в условиях постоянного обезвоживания клеток растений как механизма для предотвращения падения концентрации CO2 и истощения субстрата дыхания (путем стабилизации фотосинтеза). Быть может, этим и объясняется подавленность гликолатно-глиоксилатного пути у С4-растений и при наличии в клетках обкладки соответствующих ферментов — эти растения имеют механизм для поддержания фотосинтеза даже при Закрытых устьицах. При длительном ограничении доступа CO2 из внешней среды и у С4-растений начинает функционировать гликолатный путь.
С энергетической точки зрения фотодыхание рассматривают как «холостой» механизм, связанный с потерей энергии и уже синтезированного вещества. Однако нельзя забывать его значения для защиты окислительной системы клетки от фоторедукции в условиях высокий интенсивности света и недостатка влаги. Найден довольно эффективный способ для ослабления последствий подавления «физиологического зла». Мы здесь еще раз сталкиваемся со сложностью действия естественного отбора и самого процесса эволюции.
Энергетическая эволюция растений связана с увеличением экологического разнообразия видов и форм по эффективному использованию и сочетанию механизмов фотосинтеза и дыхания в различных условиях. Это достигалось преимущественным выживанием в критических ситуациях индивидуумов — носителей благоприятных мутаций по синтезу пигментов и набору ферментов. Поэтому энергетическая система растений развивалась постепенно, дополняя ранее возникшие механизмы новыми реакциями обмена.
Значительны различия видов растений в экологии дыхания, в чем проявляются характерные черты их приспособления. Показателем приспособленности растений в онто- и филогенезе служит и фотодыхание. Механизмы дыхания как энергетическая основа жизнедеятельности оказались базисом для возникновения последующего многообразия растений, в том числе эволюции их химического состава и ритмики.