Паратиреоидный гормон регулирует обмен кальция и поддержание постоянного уровня этого элемента в крови. При снижении уровня гормона в крови вследствие недостаточной функции паращитовидных желез или при экспериментальной паратиреоитектомии у животных развивается тяжелый болезненный синдром, называемый паратиреопривной тетанией, характеризующейся тоническими судорогами и часто заканчивающейся гибелью животных.
Химическая природа гормона окончательно не выявлена. Известно, что это белок или полипептид с молекулярной массой 9500, состоящий из 84 аминокислотных остатков. Точная последовательность аминокислот в гормоне не установлена. В настоящее время принята концепция о двояком действии паратиреоидного гормона — на почки и на кость одновременно.
Под влиянием гормона усиливается выведение фосфата с мочой вследствие либо пониженной его реабсорбции (например, у жвачных), либо усиленной секреции в дистальных участках почечных канальцев (у птиц). Наряду с этим нарастает концентрация ионов кальция в сыворотке, достигающая максимума через 12—18 ч после введения гормона. Этот эффект обусловлен непосредственным влиянием гормона на резорбцию кости с последующей мобилизацией ионов кальция.
После удаления паращитовидных желез у собак содержание кальция в сыворотке уменьшается и на третий-четвертый день падает до уровня 6—7 мг% вместо 10—11 мг% в норме, причем уменьшение идет исключительно за счет ионизированного кальция. Уровень неорганического фосфора возрастает с 3,2 до 5,7 мг%. Возбудимость нервно-мышечной системы повышается, что и является причиной развития тетании.
Установлено, что присутствие лишь одного минерального компонента кости обеспечивает за счет физико-химического равновесия поддержание уровня кальция в сыворотке крови в пределах 6—7 мг% без вмешательства паращитовидных желез. Однако благодаря функции последних минеральный компонент кости способствует более высокому содержанию кальция в плазме, чем это можно было бы ожидать, исходя из обычных законов растворимости.
Активные процессы, обусловливающие гомеостатическую регуляцию уровня кальция в крови, могут быть двоякого рода:
- поддержание кальциемии, или, точнее, активности ионизированного кальция (ACa2+), в пределах, необходимых для нормального осуществления физиологических функций;
- поддержание произведения активностей ионов кальция и фосфата ACa2+·AHPO42- — во внеклеточной жидкости на уровне, обеспечивающем спонтанный рост кристаллов гидроксиапатита в кости. Сыворотка крови пересыщена минеральными компонентами кости. Осаждению из нее трикальцийфосфата препятствует пирофосфат, содержащийся в сыворотке и тканях.
Эти два процесса необходимо различать, так как они обычно связаны обратной зависимостью. Так, при повышении фосфатемии произведение ACa2+·AHPO42- возрастает, а активность ионизированного кальция (ACa2+) снижается. Произведение активностей ACa2+·AHPO42- константно.
Паращитовидные железы непосредственно реагируют (в системе с простой отрицательной обратной связью) только на активность ионов Ca2+ в плазме и межклеточной жидкости, усиливая соответственно секрецию паратгормона. Замечено, однако, что паращитовидные железы оказывают наибольшее влияние на уровень кальция в сыворотке, когда изменяется также содержание фосфата. Приведенные выше соображения дают этому факту удовлетворительное объяснение.
Поскольку основная роль гормона состоит в восстановлении сниженной активности ионного кальция (ACa2+), а не произведения активностей (ACa2+·AHPO42-). гормон не может выполнить своей роли только за счет резорбции кости (в этом случае возможно лишь увеличение произведения активностей). Для реального повышения ACa2+ необходимо, чтобы фосфат сыворотки частично был экскретирован через почки.
Таким образом, при уменьшении количества фосфата в сыворотке за счет его повышенной почечной экскреции под влиянием паратгормона увеличивается ACa2+, что становится, по выражению Рассмуссена, «первой линией обороны» против снижения активности Са2+. Этот механизм очень быстр и чувствителен, но с ограниченными возможностями. «Второй линией обороны» против снижения ионной активности Са2+, менее чувствительной, но практически неисчерпаемой, является мобилизация ионов Са2+ из кости в результате прямого действия паратгормона. Реципрокно усиливается выведение излишка фосфатов почками.
Непосредственный механизм влияния паратгормона на резорбцию костей недостаточно ясен. Предполагается, что он связан с действием иона цитрата, выполняющего одновременно функции растворителя, хелатирующего агента и фактора, локально снижающего величину pH. Это подтверждается, в частности, относительно высоким содержанием цитрата в костной ткани и изменением его уровня в тканях организма при инъекции паратгормона или при паратиреоидектомии. Под влиянием паратгормона изменяется клеточный состав кости — уменьшается содержание остеобластов и резко возрастает число остеокластов.
Предполагается (но точно не доказано) еще два места приложения паратиреоидного гормона в организме: влияние (наряду с витамином D) на всасывание кальция в кишечнике и стимуляция реабсорбции кальция в извитых канальцах почек. Эти подсобные функции гормона усиливают конечный эффект его действия, способствуя повышению концентрации Са2+ в биологических жидкостях.
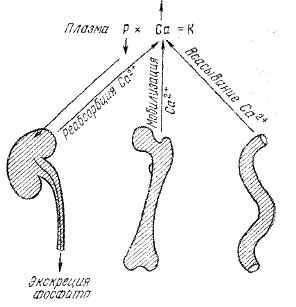
Пути действия паратгормона. Паратгормон повышает концентрацию ионов кальция в плазме. Кроме того, он снижает концентрацию фосфат-иона в плазме, компенсируя тем самым эффект высвобождения ионов фосфата при выходе кальция из кости. Благодаря этому повышение концентрации кальция в плазме происходит в соответствии с соотношением РхСа=К
Не исключено, что регулирующее влияние паратиреоидина (и кальцитонина) на абсорбцию кальция может осуществляться, путем контроля за превращением в почках витамина D в активную форму — 1,25-диоксихолекальциферол.
Классическая форма паратиреопривной тетании легко воспроизводится на собаках. У сельскохозяйственных животных она проявляется слабо, что обусловлено особенностями их питания, высокой продуктивностью, большими обменноспособными фондами кальция, возможно, наличием добавочной паратиреоидной ткани. Так, у кур-несушек операция полной или односторонней па- ратиреоидектомии не вызывает симптомов тетании, хотя и снижает кальциемию при одновременном увеличении уровня фосфата в крови. Яйцекладка при этом нарушается.
У жвачных животных клиническая картина после паратиреоидектомии (или одновременной тирео- и паратиреоидектомии) также недостаточно типична, хотя в ряде опытов наблюдалась классическая тетания. Для жвачных характерно влияние паратиреоидного гормона на обмен фосфата. После удаления желез животные меньше потребляют корма, но у них развивается не анорексия, а специфическая потеря аппетита к фосфору. Относительная величина абсорбции фосфора в кишечнике (в отличие от кальция) при этом не изменяется, но общее количество абсорбированного фосфора уменьшается. Значительно снижаются эндогенные фекальные потери фосфора (и кальция), падает уровень неорганического фосфата в крови. Введение паратиреоидного гормона интактным животным вызывает обратный эффект. По-видимому, одним из путей увеличения эндогенной экскреции фосфора является повышенное выделение его со слюной околоушных слюнных желез.