Наследственность является свойством клетки как системы в процессе ее работы и деления.
Нельзя считать, что для осуществления наследственности важно ядро и не важна цитоплазма, так как во всякой работающей системе все детали важны. Задача исследований состоит в выяснении функции и структуры отдельных компонентов клетки, которые в процессе эволюции приобрели свое специальное назначение.
Ядро можно назвать органом хранения наследственности, цитоплазму — органом осуществления наследственности.
Согласно принятому нами определению наследственности, все самовоспроизводящиеся структуры клетки должны обладать преемственностью. В этом аспекте и необходимо рассмотреть принципиальные отличия в структуре и функции ядра и цитоплазмы.
- Ядро содержит ограниченное число и характерный для каждого вида набор хромосом; цитоплазма содержит много органоидов, число которых, по-видимому, непостоянно.
- Ядро не способно исправить и заместить возникшие дефекты хромосом; поврежденные органоиды цитоплазмы могут быть замещены путем размножения в одном клеточном цикле.
- Ядро в норме обязательно репродуцируется идентично; произошедшее изменение в хромосомах воспроизводится в том же виде; органоиды цитоплазмы могут репродуцироваться количественно неидентично.
- При клеточном делении хромосомы распределяются между дочерними клетками поровну; органоиды цитоплазмы распределяются или могут распределяться неравномерно.
- Ядерная наследственность подчиняется менделевским законам наследования, а цитоплазматическая — не подчиняется.
В общей форме можно сказать, что отличительные черты в структуре и функции ядра и цитоплазмы обусловлены их специализацией и различным назначением в деятельности клетки как системы. Поскольку органы цитоплазмы, как и ядро, содержат нуклеиновые кислоты и установлена цитоплазматическая дискретная наследственность, теперь принято различать ядерные, или хромосомные, гены и внеядерные, или экстрахромосомные, гены.
Для изучения роли ядра и цитоплазмы в наследственности применяются различные методы. К ним относятся:
- метод замещения ядра;
- метод реципрокных и возвратных скрещиваний;
- метод получения цитоплазматических мутаций и изучения поведения их в поколениях.
Одним из наиболее надежных в настоящее время вариантов метода замещения ядра является разрушение ядра яйцеклетки одного вида с последующим оплодотворением ее сперматозоидом другого вида. При развитии таких зигот должны получаться гаплоидные андрогенное особи, поскольку они несут одно только отцовское ядро и материнскую цитоплазму. Но гаплоидные зиготы погибают на ранней эмбриональной или личиночной стадиях, и поэтому долгое время таким путем не удавалось получить взрослых форм андрогенного происхождения. Лишь недавно Б. Л. Астаурову и В. П. Остряковой-Варшавер удалось получить диплоидных андрогенных гибридов при скрещивании двух видов шелкопряда: Bombyx mori и B. mandarina.
У шелкопряда, как и у других бабочек, гетерогаметным полом являются самки, а гомогаметным самцы. Кроме того, у шелкопряда, как и у многих насекомых, наблюдается полиспермия. Хромосомы тутового шелкопряда легко метятся определенными генами, с которым можно различать в потомстве признаки обоих видов. В скрещивании В. mandarina X В. mori самец был маркирован тремя рецессивными генами, находящимися в разных хромосомах: ch — шоколадный, определяющий желто-коричневую окраску личинок, р — ген белой окраски бабочек и ml — ген молочно-белой окраски гиподермы гусениц. Самка была отмечена соответственно тремя доминантными аллелями этих генов, определяющими черный цвет личинок, темную окраску бабочек и серый цвет гусениц. Следовательно, при указанном скрещивании у всех нормальных гибридных потомков должны быть черные личинки, серые гусеницы и тёмные бабочки, а у андрогенных особей должны проявиться рецессивные гены: личинки желто-коричневого цвета, гусеницы и бабочки светлой окраски. В этом исследовании разрушение ядра яйцеклетки после проникновения в нее сперматозоидов производилось в момент второго мейотического деления тепловым шоком, а именно воздействием температуры 40° в течение 120—135 мин. каждой кладки одну часть яиц (грены) подвергали такому воздействию, другую — оставляли в качестве контроля. Так как в опыт-группе материнское ядро убивалось, то развитие могло идти только при условии, если сливались два мужских пронуклеуса образуя одно диплоидное ядро (диспермический андрогенез). В результате все развивающиеся особи должны быть мужского пола, так как самцы имеют две Х-хромосомы; кроме того, все потомки должны иметь рецессивные признаки, поскольку хромосомы сами были мечены рецессивными генами.
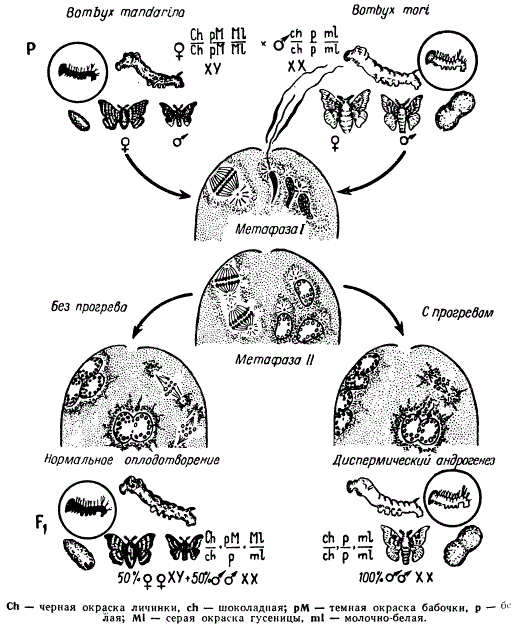
Схема получения диплоидных андрогенных особей у Bombyx методом теплового воздействия
В опыте было получено большое число андрогенных личинок и несколько половозрелых самцов, имевших сперматозоиды, способные к оплодотворению. Результаты опыта можно рассматривать как прямое доказательство ведущей роли ядра в наследственности и отсутствия заметного влияния материнской цитоплазмы.
Г. Гемерлинг провел опыт с замещением ядра у зеленой водоросли рода Acetabularia. Он взял два вида этого рода mediterranea и A. wetfsteinii, различающихся формой шляпки. Эти водоросли на определенном этапе жизненного цикла имеют по одному ядру, находящемуся в одном из ризоидов.
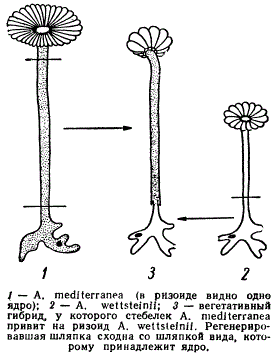
Влияние ядра на форму шляпки у Acetobularia
Отрезая ризоиды, содержащие ядро, и затем сращивая их с отрезками стебельков так, чтобы ядро одного вида соединялось с плазмой другого, можно было наблюдать влияние ядра в чужой плазме на развитие новой шляпки. Оказалось, что форма шляпки развивается соответственно тому виду, которому принадлежит пересаженное ядро.
Микрохирургическим путем удавалось замещать ядра у разных видов амеб. И в этом случае признаки вида-реципиента, сохранявшие свою цитоплазму, проявлялись лишь в первые дни после операции, а затем обнаруживались признаки вида, которому принадлежало пересаженное ядро.