Прежде чем уяснить себе строение нуклеиновых кислот, ученым пришлось тщательно исследовать их количественный состав и на основании химического анализа прийти к выводу, что существуют два различных класса макромолекул.
Оказалось, что в состав нуклеиновых кислот входят сложные кольцевые азотистые основания — производные пуринов и пиримидинов, углеводы с пятью углеродными атомами и фосфорная кислота. Вот химические формулы двух углеводов, по которым различаются два класса нуклеиновых кислот:
Нуклеиновая кислота, в состав которой входит дезоксирибоза, называется дезоксирибонуклеиновой — ДНК. В состав рибонуклеиновой кислоты, РНК, входит рибоза. ДНК и РНК различаются и по месту расположения в живой клетке — ДНК находится в ядре, а РНК в цитоплазме.
Нуклеиновые кислоты — это высокополимерные вещества, состоящие из отдельных мономеров. Звеном для построения полимерных молекул нуклеиновых кислот являются нуклеотиды. А один нуклеотид — это фосфорная кислота и углевод, соединенные с пуриновым или пиримидиновым основанием.
РНК отличается от ДНК еще и тем, что вместо тимина, содержащегося в ДНК, в ее состав входит урацил.
Установлению состава нуклеиновых кислот предшествовало тщательное его изучение у различных представителей живого мира — вирусов, бактерий, растений и у более высокоорганизованных существ. Исследование нуклеотидного состава нуклеиновых кислот позволило выяснить одну важную деталь.
Оказалось, что каждый вид организма (особенно ярко это было выражено у бактерий) характеризуется специфическим типом ДНК. Химический анализ ДНК, проведенный американцем Чаргаффом, позволил ему открыть следующие правила.
Во-первых, сумма пуриновых оснований, аденина и гуанина, А и Г, равна сумме пиримидиновых, цитозина и тимина и, во-вторых, количество аденина всегда равно содержанию тимина, а количество гуанина — числу оснований цитозина. Эта важнейшая особенность в химическом строении нуклеиновых кислот находит отражение в так называемом показателе специфичности ДНК: (Г+Ц):(А+Т). Нуклеотидный состав и показатель специфичности ДИК различных биологических видов приведен в таблице.
Очень большое значение в установлении указанных закономерностей имели исследования нуклеотидного состава ДНК у различных представителей животного и растительного мира, выполненные школой советских биохимиков, во главе которой стоит академик А. Н. Белозерский.
Параллельно с изучением химического состава и соотношения оснований в ДНК различных биологических видов ученые исследовали и структуру ДНК. Применив весь арсенал методов, принятых в экспериментах с высокополимерными веществами, ученые обнаружили, что ДНК имеет очень большой молекулярный вес. Если молекулярный вес только некоторых белков достигает нескольких миллионов, то вес ДНК, выделенной из различных биологических источников, равен 6—10 миллионам. Сейчас часть исследователей считает, что молекулы ДНК размером в 6—10 миллионов — это осколки еще больших молекул, составляющих ядро клетки, которые могут повреждаться в момент разрушения ядра при выделении нуклеиновых кислот.
Это мнение далеко не беспочвенно, так как недавно из бактериофага удалось выделить ДНК с молекулярным весом в 130 миллионов. Чтобы получить представление о размерах такой молекулы, достаточно сказать, что ее длина равна 70 микронам. А из кишечной палочки недавно выделена молекула ДНК длиной почти в сантиметр. Поистине, сантиметровые молекулы!
Как и белки, нуклеиновые кислоты являются линейными полимерами. По аналогии с белковыми молекулами первичная структура нуклеиновых кислот — это линейное расположение нуклеотидов один за другим. Но наиболее интересная особенность строения ДНК — ее вторичная структура.
В первые годы после второй мировой войны в Англии начались интенсивные исследования ДНК с помощью рентгеноструктурного анализа. Одно из требований, которое необходимо выполнять при изучении строения веществ с помощью рентгеноструктурного анализа — обязательное предварительное ориентирование полимерных молекул. Это легко понять, если представить себе высокополимерную молекулу в виде спирально загнутой проволоки — пружины. Если диаметр пружины очень мал, то частоту завивки (шаг спирали) можно рассмотреть при проектировании на экран увеличенного изображения с помощью источника света. Одну молекулу — пружину взять невозможно, так как она слишком мала для этого. Если же взять беспорядочный клубок молекул, то детали спирализации увидеть будет очень трудно. Поэтому ученые приготовили ДНК в виде тонкой нити, состоящей из упакованных по длине молекул, находящихся в ориентированном состоянии.
Первые рентгенограммы, которые удалось получить Уилкинсу и Франклин, содержали очень большое количество пятен. Математическая обработка этих рентгенограмм позволила ученым выяснить некоторые основные принципы строения молекул ДНК. Регулярность расположения пятен на снимках указывала на спиральное строение ДНК, с длиной одного витка 34 Å, причем, по-видимому, спираль составляли две полимерные молекулы. Эта сухая информация отнюдь не была исчерпывающей для понимания структуры ДНК, но остальное сделала блестящая интуиция ученых. Однако догадка вовсе не была беспочвенной, так как она базировалась на богатейшем аналитическом материале, из которого было получено правило Чаргаффа, и на знании пространственной конфигурации нуклеотидов — элементарных ячеек, из которых состоят нуклеиновые кислоты.
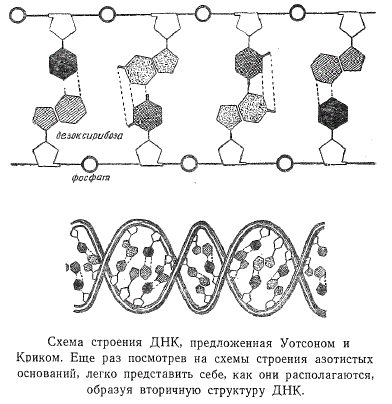
Американский ученый Уотсон и англичанин Крик высказали предположение, что каждая из двух спиралей — это соединенные между собой в единую непрерывную цепь дезоксирибоза и фосфатный остаток. Азотистые основания каждой из двух спиралей располагаются перпендикулярно к ее основному направлению и соединяются с основаниями противоположной спирали по правилу Чаргаффа. Напомним, что из анализа нуклеотидного состава ДНК следует, что количество молекул аденина всегда равно количеству тимина, а содержание гуанина эквивалентно таковому цитозина. Иными словами, аденин всегда соединяется с тимином, а гуанин с цитозином. Такая схема строения ДНК хорошо соответствовала условиям, которые предъявлялись на основании исследования пространственной картины строения азотистых оснований.
Представьте себе монету, разрезанную на две неравные части, и полую трубку, внутренний диаметр которой равен диаметру монеты. Предположим, что меньшая половина — это пиримидиновые основания, большая — пуриновые, а толщина молекулы ДНК равна диаметру полой трубки. Ясно, что сложив только пуриновое и пиримидиновое основание, можно получить в результате целый монетный кружок, который точно закроет трубку. Комбинация пуринового основания с себе подобным приведет к образованию большего кружка, а пиримидинового с пиримидиновым — меньшего. Таким образом, в схеме Уотсона — Крика учитывалось условие пространственного соответствия, по которому сумма расстояний между боковыми группами оснований пуринов и пиримидинов давала постоянную величину.
Необходимо сказать в дополнение несколько слов о направлении, в котором закручены обе спирали. Они антипараллельны, то есть завернуты в противоположном направлении вокруг общей оси, как это показано на рисунке.
Азотистые основания располагаются друг над другом подобно стопке монет, а расстояние между плоскостями соседних пар равно 3,4 Å. Пары аденин — тимин и гуанин — цитозин связаны между собой так называемыми водородными связями, которые осуществляются через атом водорода.
Эта схема сейчас общепризнана, а ее авторы Уотсон и Крик и пионер рентгеноструктурных исследований ДНК англичанин Уилкинс в 1962 году были удостоены Нобелевской премии.
Модель строения ДНК заключает в себе очень глубокий биологический смысл. Так как аденин соединяется только с тимином, а гуанин с цитозином, то каждая спираль является зеркальным отражением другой спирали, то есть точным штампом для ее построения.
Открытие строения молекулы ДНК явилось первым шагом к попытке объяснить на молекулярном уровне механизмы наследственности и биологического синтеза ДНК.