Наличие наследственной изменчивости позволяет путем различных систем скрещивания сочетать определенные наследственные признаки в одном организме, а также избавляться от нежелательных свойств.
Основным методом управления комбинативной изменчивостью в селекции является подбор форм по хозяйственно ценным свойствам для скрещивания.
Классификация типов скрещивания и методов разведения
При разведении применяют различные системы скрещиваний, которые понято делить на родственное скрещивание, часто называемое инбридингом, или инцухтом, и неродственное, иногда называемое аутбридингом.
Разновидностью аутбридинга является межпородное скрещивание (кроссбридинг). Инбридинг — английский термин, в русской литературе чаще употребляется для обозначения близкородственного разведения у животных, инцухт — немецкий термин, употребляется для обозначения принудительного самоопыления у перекрестноопыляющихся растений. Однако во избежание путаницы можно пользоваться одним термином — инбридинг.
В животноводстве скрещивания делят на два типа в соответствии с задачей разведения животных: племенное (заводское) и промышленное (товарное). Для собственно селекционных целей и племенного разведения, т. е выведения новых пород и улучшения породных свойств, применяется как инбридинг, так и аутбридинг. Для увеличения продуктивности животноводства используют промышленное скрещивание на основе существующих пород. Подобные же типы скрещиваний применяются и в современной селекции растений для выведения или поддержания сорта и для получения товарного продукта. Например, скрещивания, применяемые у сахарной свеклы или арбуза для получения триплоидных семян, являются типично промышленными.
Применение той или иной системы скрещиваний в селекции зависит от того, какой тип изменчивости используется для селекционных целей и какие задачи при этом решаются. Например, если скрещивание подобранных форм (комбинативная изменчивость) не дает эффекта, то прибегают к использованию мутационной изменчивости или к полиплоидии. При этом изменяют и систему скрещиваний.
Подбор для скрещивания исходных форм производят из популяций. Для правильного выбора исходных форм необходимо сначала оценить генетический состав (потенциал) популяции, из которой они происходят. Так, очевидно, что для получения жирномолочных коров необходимо скрещивать животных, происходящих из популяции, имеющей высокую концентрацию генов жирномолочности, а для получения овец с мериносовой шерстью необходимо скрещивать животных из популяции тонкорунных, а не грубошерстных овец.
Изучение генетического состава исходных популяций и их происхождения облегчает создание соответствующих генотипов. Таким образом, оценка исходных популяций животных является первейшей задачей селекции, которая должна осуществляться селекционными и генетическими методами на основе анализа различных показателей продуктивности.
Родственное скрещивание
Генетический состав популяции оценивается путем разложения ее на генетически различные линии.
Для автогамных организмов, как показал В. Иоганнсен, это достигается просто — путем выделения потомства отдельных самоопыляющихся растений, а для аллогамных организмов необходимо проводить инбридинг.
Родственным называют скрещивание особей, имеющих близкую степень родства: брат — сестра, отец — дочь, мать — сын, двоюродные братья и сестры и т. д. Разную степень родства скрещиваемых животных, т. е. большее или меньшее сходство их генотипов, определяют с помощью коэффициента генетического родства. У растений наиболее тесная форма инбридинга осуществляется при принудительном самоопылении.
Генетическая сущность инбридинга сводится к процессу разложения популяции на линии с различными генотипами. При этом гены, находящиеся в гетерозиготном состоянии, переходят в гомозиготное состояние. Например, при скрещивании самца и самки, гетерозиготных по одному гену (Аа), в потомстве будет расщепление 1АА : 2Аа : 1аа, или в процентном отношении 25АА : 50Аа и 25аа. Если в ряду последующих поколений каждый из генотипов будет скрещиваться внутри себя, т. е. будет осуществляться инбридинг, то в последующих поколениях число гомозиготных форм увеличится, а гетерозиготных — сократится.
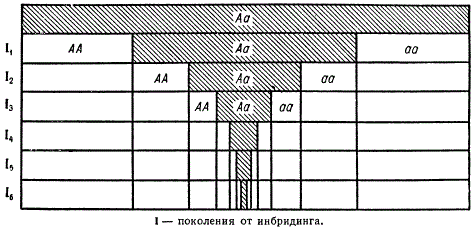
Движение популяции при инбридинге (динамика соотношения гомо- и гетерозигот яри расщеплении по одной паре аллелей)
Представим теперь, что аллель а имеет летальное действие, т. е. резко снижает жизнеспособность. Очевидно, что в каждом поколении инбридинга 25% особей (аа) будут либо отмирать, либо проявят пониженную жизнеспособность. В результате инбридинг в ряду поколений приведет к депрессии.
В связи с тем, что каждый перекрестноопыляющийся сорт, как мы можем убедиться на примере разных сортов кукурузы, насыщен различными вредными рецессивными мутациями, то естественно, что при инбридинге часто проявляется понижение жизнеспособности, урожайности, устойчивости к заболеваниям и т. п. Доказательством этого могут служить данные Д. Джонса по влиянию инбридинга в течение 15 поколений на урожайность зерна и высоту растений в четырех линиях кукурузы А, В, С и D. Приведенные данные показывают, что исходные формы фенотипически были одинаковы. Применение принудительного самоопыления во всех линиях привело к снижению урожайности и высоты растений. При этом в одних линиях депрессия наступила скорее, чем в других. Это может указывать на то, что гомозиготность по рецессивным генам наступает в разных линиях с разной скоростью. Последняя зависит от многих факторов: от числа генов, по которым имелась гетерозиготность, от степени родства скрещиваемых форм и др.
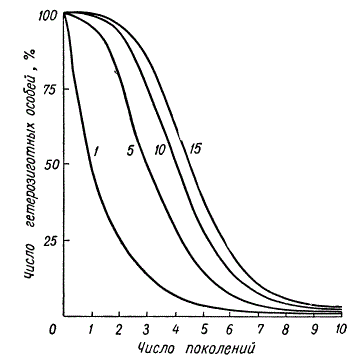
Гомозиготизация в инбредной популяции в зависимости от числа пар аллелей (1, 5, 10, 15), определяющих признак.
На рисунке выше показано сокращение процента гетерозиготных особей в различных поколениях инбридинга в зависимости от числа генов, по которым имелась гетерозиготность. Чем большее число разных генов определяет какой-то признак или свойство, тем медленнее наступает гомозиготное состояние по всем рецессивным аллелям, тем медленнее идет стабилизация признака. Уменьшение процента гетерозиготных особей в последовательных поколениях инбридинга в зависимости от степени родства скрещиваемых особей показано на рисунке ниже.
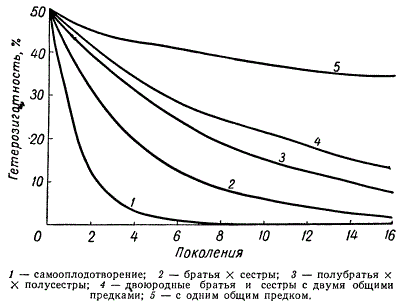
Встречаемость гетерозиготных особей в популяции в зависимости от степени родства скрещиваемых форм
Гомозиготность наступает наиболее быстро при самооплодотворении. При системе скрещивания «брат X сестра» процент гетерозиготных особей в ряду поколений снижается медленнее, но все же быстрее, чем при скрещивании двоюродных братьев и сестер или при еще более дальнем родстве скрещиваемых организмов.
Все эти расчеты справедливы лишь для случаев, когда гены находятся в разных негомологичных хромосомах. На самом деле гены, определяющие одни и те же свойства, могут находиться в одной группе сцепления на разном расстоянии друг от друга и претерпевать различную частоту перекреста. Кроме того, в этих расчетах не учитывается мутационная изменчивость генов, взаимодействие генов в системе генотипа и, самое главное, действие искусственного и естественного отбора, зачастую благоприятствующего сохранению гетерозиготных форм. Но, несмотря на формальный характер подобных расчетов, они дают возможность правильно выбрать систему скрещивания для наследственного закрепления свойств в выводимой породе или сорте.
Относительно полезности и вредности применения инбридинга в селекции существуют различные суждения. Действительно, при применении инбридинга у животных и аллогамных растений (кукуруза, рожь и другие) очень быстро наступает депрессия по жизнеспособности, плодовитости и другим свойствам. Если в стаде кур ежегодно получать потомство путем спаривания «брат х сестра», то в течение нескольких поколений заметно снижается яйценоскость и жизнеспособность кур, чаще появляются различные уродства. Такое же явление наблюдается при инбридинге у свиней и многих других животных. На этом же основании существует запрещение родственных браков в человеческом обществе.
Однако известно, что в природе существуют виды растений и животных, для которых автогамное размножение является нормой, и при этом они не только не вымирают, а, наоборот, процветают. К таким растениям относятся ячмень, пшеница, горох, фасоль и др. Оказывается, самоопыление и самооплодотворение не приводят к депрессии те виды, у которых этот процесс приобрел приспособительное значение для наиболее надежного обеспечения размножения.
Каким же образом объяснить тот факт, что инбридинг может быть и полезным, и вредным?
В процессе инбридинга депрессию вызывают мутантные аллели, понижающие жизнеспособность организмов. В гетерозиготном состоянии их действие подавляется доминантными, нормальными, аллелями. Поэтому при свободном скрещивании в популяции они не выявляются с такой частотой, как при инбридинге. Но среди мутаций могут быть не только вредные, понижающие жизнеспособность, но и повышающие ее, особенно при благоприятном сочетании генов. Отсюда следует, что не всегда при близкородственном размножении животных или растений может наступать депрессия. Напротив, могут выделиться линии с повышенной жизнеспособностью и продуктивностью. Но это происходит в чрезвычайно редких случаях, так как число вредных рецессивных мутаций значительно превышает число полезных. Этим можно объяснить приспособительное значение гетерозиготности у перекрестнооплодотворяющихся организмов и самого перекрестного опыления и скрещивания. Следовательно, вреден не сам по себе инбридинг, а последствия гомозиготизации вредных мутаций и снижение оптимального уровня гетерозиготности популяции. При умелом применении инбридинга удается выделить ценные генотипы.
Подобно тому как луч света, прошедший через призму, разлагается на целый спектр хроматических линий, так и популяция гетерозиготных организмов с помощью инбридинга может быть разложена на отдельные, генетически различающиеся линии. Инбридинг позволяет выделить из популяции группы организмов с отдельными необходимыми для селекции свойствами. В «кровной линии», в которой родственные по происхождению организмы скрещиваются между собой, повышается концентрация отдельных генов, вследствие чего внутри линии увеличивается число гомозиготных особей. Поэтому особи внутри каждой линии оказываются менее изменчивы, более однородны и надежнее передают свои свойства потомству. Линия, которую часто называют инбридированной, или инбредной, в меньшей степени расщепляется на разные генотипы.
Встает вопрос: можно ли при длительном инбридинге получить абсолютно гомозиготные формы? Опираясь на знания генетики, на этот вопрос следует ответить отрицательно. Во-первых, естественный отбор, как правило, поддерживает оптимальный уровень гетерозиготности; во-вторых, наличие сцепления и перекреста хромосом в значительной степени задерживает гомозиготизацию в ряду многих поколений инбридинга и также может давать новые комбинации генов в генотипах потомков; в-третьих, непрерывно возникает множество различных мутаций, которые будут нарушать гомозиготность линий; мутация даже одного гена может привести к изменению генотипической нормы реакции всего организма.
В силу указанных причин линии, полученные при длительном инбридинге, имеют лишь относительную гомозиготность. Благодаря этому и в таких линиях отбор может давать некоторый эффект. Очевидно, что на первых этапах инбридинга отбор может давать гораздо более значительный сдвиг в желаемую сторону, чем в последующих поколениях. Отбор при высоких степенях инбридинга менее эффективен, но зато повышается гарантия наследственного закрепления отбираемых свойств.
Неродственное скрещивание (разведение)
Прямой противоположностью инбридингу служит скрещивание неродственных по происхождению организмов, или аутбридинг.
Конечно, все организмы, относящиеся к одному виду или роду, имеют общее происхождение. Но когда мы говорим о неродственном скрещивании, то имеется в виду отсутствие у скрещиваемых особей ближайших общих предков в 4—6 поколениях их родословных (прадеда, деда, прабабушки, бабушки и т. п.). Чаще неродственным скрещиванием организмов называют такое, при котором родительские формы происходят из разных генетических популяций.
При скрещивании неродственных особей вредные рецессивные мутанты, находящиеся в гомозиготном состоянии, перейдут в гетерозиготное состояние и не будут оказывать влияния на жизнеспособность гибридного организма. Действительно, весь опыт практики сельского хозяйства показывает, что скрещивание неродственных организмов внутри одного и того же вида чаще ведет к тому, что помеси первого поколения оказываются более жизнеспособными, более устойчивыми к заболеваниям, имеют повышенную плодовитость, т. е. проявляют гетерозис.
Неродственное скрещивание служит важным методом селекции и разведения. Путем этого скрещивания производят объединение разных наследственных свойств в одном гибридном организме. С его помощью комбинируют различные ценные признаки для создания новой породы или сорта. Так, например, для того чтобы повысить живой вес кур породы леггорн, их можно скрестить с петухом другой породы, характеризующейся большим живым весом, например с белым плимутроком. Гибридные куры первого поколения по весу будут занимать промежуточное положение и окажутся в среднем большего веса, чем леггорны. Но если их скрестить с такими же гибридными петухами, то во втором поколении произойдет расщепление на различные по весу особи. Породы еще не будет, но зато в этом поколении могут встретиться нужные нам сочетания признаков. Дело селекционера — отобрать наиболее ценные генотипы. При этом отбор, как мы увидим дальше, должен производиться только по фенотипу, но и по генотипу.
Из сказанного следует твердо усвоить, что при аутбридинге первое поколение по сложным наследственным признакам, как правило, будет промежуточным и более единообразным, чем второе поколение, так как в последнем происходит расщепление. И если в последующем не будут применены определенная система разведения и строгий отбор, то новой породы создать не удастся, а исходные утратят свою породность. То же самое относится к скрещиваниям разных пород крупного и мелкого рогатого скота и свиней, а также к сортам растений.