После установления групп сцепления генов и составления генетических карт на основе учета перекреста хромосом возникла необходимость составления цитологических карт с целью их сопоставления с генетическими. Это было осуществлено сначала на митотических, а затем на гигантских хромосомах.
Один из методов основывался на использовании явления хромосомных перестроек, в частности транслокаций. Сущность хромосомных перестроек типа транслокаций заключается в следующем: если в двух негомологичных хромосомах одновременно происходит разрыв, то фрагменты этих негомологичных хромосом могут обменяться местами. Поскольку каждый фрагмент с генами присоединяется к новой для него группе сцепления, то можно установить в обеих хромосомах точки разлома и присоединения фрагментов к негомологичным хромосомам, а также измерить величину фрагментов. Устанавливается это генетически, с помощью кроссинговера, и цитологически, в световом микроскопе путем непосредственного измерения переместившихся фрагментов на пахитенной стадии мейоза.
Маркируя хромосому различными генами и зная место их нахождения на генетической карте, можно по месту разрыва, произошедшего вблизи известного гена, определить относительное расстояние между генами в самой хромосоме. Таким методом Ф. Г. Добржанскому удалось составить первые цитологические карты хромосом и сопоставить их с генетическими.
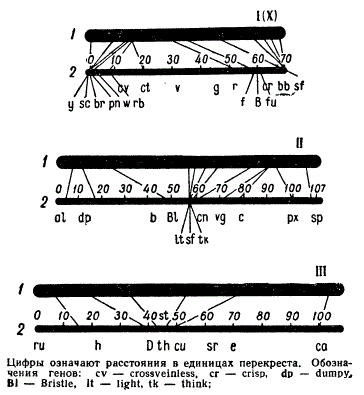
Сравнение относительных размеров соответствующих участков на цитологических (1) и генетических (2) картах хромосом (I, II, III) Drosophila.
Эти цитологические карты полностью подтвердили ту последовательность расположения генов, которая была установлена на основе чисто генетических методов. Несовпадений между генетическими и цитологическими картами обнаруживалось лишь в величине расстояний между генами, причем в одних участках хромосом эти расстояния оказались меньше, в других — больше. Это объясняется тем, что в разных районах хромосом вероятность осуществления перекреста неодинакова, а частота перекреста зависит не только от расстояния между генами, но и от местонахождения района хромосомы, в котором он находится.
Цитогенетическое изучение гигантских хромосом в слюнных железах личинок дрозофилы позволило уточнить место целого ряда генов в хромосоме. Так например, на рисунке дана цитологическая карта одной из четырех гигантских хромосом дрозофилы, а именно четвертой, самой маленькой, хромосомы с указанием районов, в которых локализованы гены.
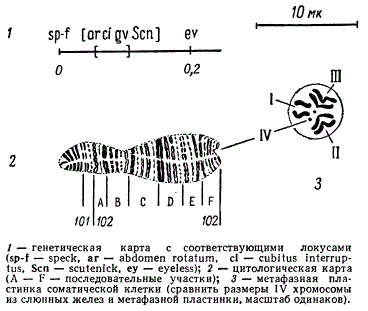
Цитологическая и генетическая карты IV хромосомы клеток слюнных желез Drosophila
Локализация генов в определенных участках хромосомы производится по методу Т. С. Пайнтера с помощью различных хромосомных перестроек: удвоения, перехватки отдельных дисков и др.
Сопоставление генетических и цитологических карт позволило скрыть еще одно явление, а именно неравномерность частоты перекрёста по длине хромосом. Это было показано на хромосомах слюнных желез следующим образом. Генетические карты всех четырех хромосом дрозофилы, составленные на основе частоты перекреста, имеют определенную длину, выражаемую в единицах перекреста. Общая длина Х-хромосомы и трех аутосом по генетическим картам составляет 279 морганид перекреста. К. Бриджес изменял длину каждой из четырех хромосом слюнных желез в микронах. Общая физическая длина хромосом слюнных желез, измеренных под микроскопом, составляет 1180 мк.
Чтобы сопоставить цитологические карты гигантских хромосом с генетическими картами для каждой хромосомы, К. Бриджес предложил воспользоваться коэффициентом кроссинговера. Для этого он разделил общую длину всех хромосом слюнных желез (1180 мк) на общую длину генетических картах (279 единиц). В среднем это отношение оказалось равным 4,2. Следовательно, каждой единице перекреста на генетической карте соответствует 4,2 мк на цитологической карте (для хромосом слюнных желез). Зная расстояние между двумя генами по генетической карте, например геном рх (с локусом 100,5) и sp (с локусом 107,0) в правом плече второй хромосомы, можно сравнить частоту перекреста в разных районах хромосомы.
В силу неравномерного осуществления обменов по длине хромосом гены при локализации, ведущейся по частоте рекомбинантов, распределяются на генетической карте с разной плотностью. Следовательно, распределение генов на генетических картах можно рассматривать как функцию распределения возможности осуществления перекреста по длине хромосомы.
Таким образом, определение местоположения генов на генетических и цитологических картах на примере дрозофилы и других объектов производилось:
- цитологически — на хромосомах в мейозе с использованием хромосомных перестроек;
- цитологически — на хромосомах слюнных желез (у дрозофилы);
- генетически — при учете мейотического кроссинговера по проценту рекомбинантных особей (зигот) у диплоидных организмов и гамет (или спор) у гаплоидных организмов;
- генетически — с учетом соматического (митотического) кроссинговера.
Сопоставление генетических и цитологических карт дало возможность подтвердить следующие положения хромосомной теории наследственности:
- хромосомы по своей длине наследственно дискретны;
- каждый ген приурочен к определенному месту — локусу в хромосоме;
- гены распределены в хромосоме в определенной линейной последовательности;
- частота кроссинговера между генами является функцией цитологического расстояния между генами.