Хромосомы животных и растений, обнаруживаемые на стадии метафазы, малы по размерам (несколько микрон), компактны и не всегда доступны для тонкого анализа, выявления их структурной дискретности.
Цитологи открыли в некоторых соматических клетках особый тип хромосом — так называемые гигантские хромосомы. Гигантские хромосомы в 100—200 раз длиннее и в 1000 раз больше содержат хромонем, чем обычные метафазные хромосомы большинства соматических и половых клеток.
Гигантские хромосомы были обнаружены итальянским цитологом Е. Бальбиани в 1881 г. в слюнных железах личинок хирономуса (Chironomus). В дальнейшем оказалось, что такая структура хромосом характерна для ядер ряда соматических клеток личинок двукрылых — для клеток кишечника, мальпигиевых сосудов, слюнных желез, а также найдена у некоторых растений в синергидах и у простейших. Наиболее типичные гигантские хромосомы можно наблюдать в слюнных железах личинок дрозофилы.
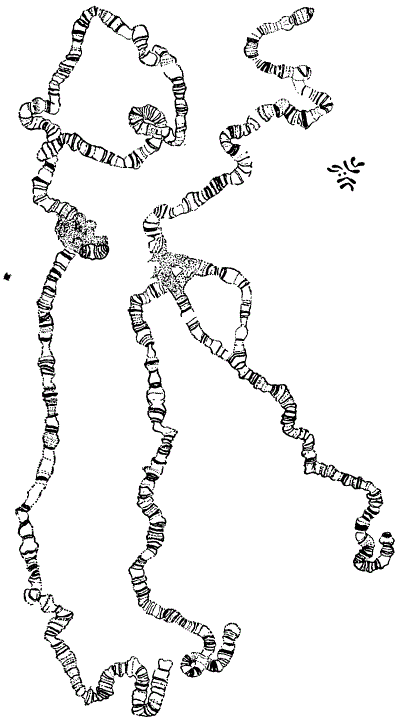
Относительные размеры гигантских хромосом ядер слюнных желез и метафазных хромосом в соматических клетках Drosophila
Гигантские хромосомы в слюнных железах личинок двукрылых возникают потому, что их клетки не претерпевают деления, а лишь увеличиваются в размерах. Ядра в течение всего личиночного периода находятся в интерфазном состоянии, а парные хромосомы (гомологи) взаимно притягиваются, вследствие чего они представляются в виде перевитых двойных тяжей.
Взаимопритяжение гомологов в интерфазных ядрах соматических клеток слюнных желез является исключением из правила и получило название соматической конъюгации хромосом. По мере роста личинки каждая из хромосом увеличивается в диаметре. Так как хромонемы гигантской хромосомы репродуцируются без последующего расхождения, то каждая хромосома приобретает вид пучка хромонемных нитей, поэтому гигантские хромосомы и называются политенными. В данном случае политения хромосом осуществляется за счет эндомитоза, т. е. редупликации хромосом без их расхождения.
Гигантские хромосомы обладают рядом морфологических особенностей. Каждая гигантская хромосома состоит более чем из 1000 хромонем, и на препаратах можно видеть отдельные нити. Предполагают, что они находятся в состоянии спирализации, которая неравномерна по оси хромосомы. Хромонемы неоднородны вдоль оси также химически и структурно, что придает нити четкообразный вид. При редупликации хромонем на хромосомах выявляются утолщения (диски), обусловленные либо более плотной локальной спирализацией, либо наличием гранул в определенных участках. Возможно, что существуют и другие причины, вызывающие различие в толщине и морфологии дисков. Диски могут быть различного размера и строения.
При конъюгации гомологов в норме идентичные диски оказываются друг против друга. Расположение этих дисков, имеющих различную морфологию, строго постоянно. Их характер и расположение можно точно устанавливать в различных клетках и у разных организмов на определенных стадиях развития. У каждого вида имеется свой «рисунок» расположения и формы дисков, по которому каждую пару хромосом можно точно находить на препаратах.
Одни авторы предполагают, что темноокрашивающиеся диски представляют собой скопление хроматина (несут в основном ДНК), а неокрашивающиеся содержат большее количество белка. Другие авторы полагают, что диски представляют собой участки хромонем с более плотной спирализацией. В таком случае темные диски должны иметь химическую природу, сходную с неокрашенными участками. Вероятно, последняя точка зрения более оправдана.
Главным результатом изучения гигантских хромосом явилось то, что расположение отдельных генов, установленное чисто генетическим методом, удалось связать с определенными районами гигантских хромосом. Некоторые гены оказались приуроченными к определенным дискам. Однако сами диски не представляют собой генов.
Разработка цитогенетического метода исследования материальных структур наследственности послужила важным этапом исследования генетической и цитологической дискретности хромосом. Хотя отдельные диски хромосом, конечно, нельзя идентифицировать с генами, которые, как мы увидим дальше, являются выражением скорее химической дискретности на молекулярном уровне, чем морфологической, тем не менее они являются важными морфологическими маркерами хромосомы, облегчающими исследование тонкого строения и функции материальных носителей наследственности.
С открытием гигантских хромосом цитогенетика значительно расширила возможности цитологического контроля различных генетических процессов.
К другому типу гигантских хромосом относят хромосомы, названные «ламповыми щетками» из-за расположения конъюгирующих хромосом в одном комплексе, напоминающем по виду ламповую щетку, или ершик.
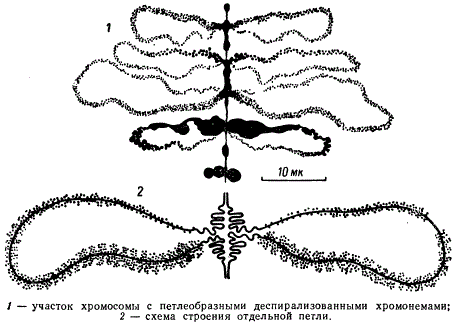
Хромосомы типа «ламповых щёток» в ооците I порядка
Эти хромосомы сильно вытянуты и образуют симметричные петли, перпендикулярные оси комплекса. Такое состояние хромосом встречается в ооцитах амфибий, рыб, и рептилий. Самые крупные хромосомы подобного типа, достигающие 700—800 ммк найдены в ооцитах саламандры.
Каждая пара гомологичных хромосом, состоящая из 4 нитей — хромонем, по оси образует серию хромомер, из которых выходят белковые петли. Полагают, что через хромомеры и петли проходит
непрерывная молекула ДНК. Хромомеры содержат ДНК, связанную с гистонами. Боковые петли наряду с ДНК содержат белки и РНК. По данным Г. Каллана, обработка «ламповых щеток» ферментом дезоксирибонуклеазой, разрывающей молекулу ДНК, показала, что хромомеры содержат 4 нити, а петли — 2 нити ДНК. На боковых петлях синтезируются РНК и белки.
В отличие от гигантских хромосом двукрылых «ламповые щетки» не являются политенными, а содержат сильно деспирализованные хромонемы. Предполагают, что большая степень деспирализации связана с повышением метаболической активности хромосом в процессе роста ооцитов.