Функции минеральных элементов в организме чрезвычайно многообразны и неразрывно связаны с их формой и состоянием. Основные из них следующие: участие в построении опорных тканей организма; поддержание гомеостаза внутренней среды; поддержание равновесия клеточных мембран; активация биохимических реакций путем воздействия на ферментные системы: прямое или косвенное влияние на функцию эндокринных желез; воздействие на симбиотическую микрофлору желудочно-кишечного тракта.
Рост животных неразрывно связан с отложением в теле минеральных веществ. Последние входят в состав всех структурных образований организма, но главным образом костной ткани, где они находятся в виде прочных нерастворимых соединений. В костях скелета сосредоточено более 80% неорганических солей организма.
Костная ткань состоит из трех основных компонентов: органической основы, неорганической фракции (костной золы) и воды, между которыми существует тесная структурная связь. В зрелой кости эти компоненты находятся в следующем примерном процентном соотношении: 38:32:30. Это соотношение зависит от возраста животного, условий его кормления и содержания. Оно неодинаково в разных костях скелета, в разных участках одной кости и даже в разных гистологических структурах. Органическая фракция кости на 95% представлена коллагеном, структура которого типична для волокнистой соединительной ткани. Коллагеновые волокна связаны гелеобразным мукополисахаридом, индентифицированным как хондроитинсульфат.
Кристаллы минерального компонента кости образованы двумя — четырьмя элементарными ячейками гидрооксиапатита, каждая из которых имеет формулу Ca10(PO4)6(OH)2. По форме кристаллы представляют собой гексагональные пластинки величиной 350x300x50 ангстрем; их поверхность на 1 г апатита достигает 100—300 м2, в связи с чем общая поверхность микрокристаллов кости необычайно велика (0,7—1,0 км2).
Существовавшее до недавнего времени мнение об инертности кости, ее неспособности к структурным изменениям, в настоящее время благодаря внедрению новых методов исследования сменилось представлением о высокой реактивности и пластичности костной ткани, ее способности к постоянной перестройке. Процесс перестройки (включая новообразование кристаллов на вновь синтезируемом матриксе и резорбции кости) происходит в течение всей жизни животного, однако в большей степени проявляется в скелете молодого растущего организма. С возрастом рост и перестройка костной ткани замедляются, что приводит у взрослых организмов к увеличению доли неактивной и полностью минерализованной костной ткани.
Механизм минерализации костной ткани изучен еще недостаточно, однако имеющиеся сведения позволяют описать его в общих чертах следующим образом. Сначала синтезируется органический матрикс — коллаген, мукополисахариды, белково-углеводные комплексы и АТФ. В этой фазе среда обогащается ионами кальция и фосфата, адсорбированными мукополисахаридами без образования ядер кристаллизации (мукополисахариды обладают повышенным сродством к ионам кальция и фосфата). Перед началом отложения солей в зону минерализации с кровью доставляется кислород, усиливаются процессы окисления и образования АТФ. Повышается активность остеобластов, выделяющих ферменты, которые влияют на основное вещество и на белковоуглеводные комплексы. В коллагеновых волокнах освобождаются реакционноспособные группы — специфически взаиморасположенные боковые аминокислотные цепи, способные служить центрами кристаллизации. Это является следствием деполимеризации части кислых мукополисахаридов, ранее блокировавших реактивные аминогруппы.
Освобождающиеся реактивные группы коллагена взаимодействуют с фосфатами и кальцием, ранее связанными мукополисахаридами, и образуют ядра кристаллизации.
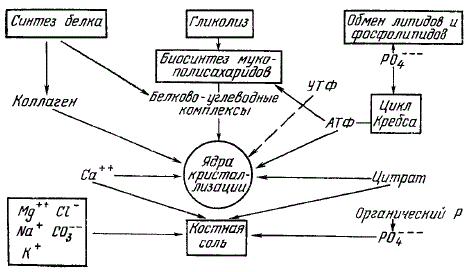
Метаболические реакции при минерализации кости
Дальнейшему осаждению кристаллов способствует то обстоятельство, что нормальная сыворотка недонасыщена по отношению к вторичному фосфату кальция (что исключает его осаждение в сыворотке), но пересыщена по отношению к гидроксиапатиту. В результате этого в присутствии костных кристаллов ионы из жидкой фазы в плотную осаждаются спонтанно. Таким образом, общепринятой в настоящее время является гипотеза эпитаксии, т. е. ориентированного роста кристаллов в центрах кристаллизации при активирующем воздействии коллагеновой матрицы.
Фермент щелочная фосфатаза, согласно этой гипотезе, принимает непосредственное участие в образовании коллагена и способствует переносу иона РО43- от эфиров к органическому основанию костной ткани:
![]()
Содержание щелочной фосфатазы пропорционально активности остеобластов. Высокий ее уровень наблюдается во время интенсивного образования коллагена и перед началом его обызвествления. Кислая фосфатаза, наоборот, сосредоточена в остеокластах и принимает непосредственное участие в процессе резорбции костей.
Отсутствие минерализации в некальцифицируемых хрящах и коллагеносодержащих тканях объясняется либо отсутствием фактора, индуцирующего кальцификацию, либо наличием факторов — ингибиторов. Последними, возможно, являются пирофосфаты и мукополисахариды, гидролизующие соответствующие энзимы (фосфатазы или сульфатазы). Вместе с тем специфическая АТФ-аза, содержащаяся в хряще, ответственна за механизм его обызвествления.
Что касается резорбции кости (т. е. одновременного рассасывания органического и неорганического компонентов), то она наряду с новообразованием костной ткани происходит в течение всей жизни животного.
Особое значение процессы резорбции приобретают у животных в период напряженного минерального обмена (при беременности, лактации, яйцекладке). В этом случае скелет выполняет функцию лабильного депо минеральных веществ, которые расходуются для нужд плода и для образования продукции (молока, шерсти, скорлупы яиц и т. д.).
Поверхность микрокристаллов оксиапатита достигает огромных размеров, и на ней могут происходить реакции замещения ионов кальция или фосфата ионами, содержащимися в жидкостях организма. Поэтому минеральный компонент кости не является чистым оксиапатитом, а содержит примеси карбоната (6%), цитрата (1%), натрия (0,7%), магния (0,7%) и следы фтора.
Значительное содержание в оксиапатите воды свидетельствует о замещении кальция в кристаллической решетке ионами (H3O)—. Кроме того, ионы гидроксония могут вытеснять ионы кальция и с поверхности кристаллов.
Общий состав минерального компонента кости с учетом молярных соотношений всех примесей можно выразить формулой:
![]()
Согласно Ньюману и Ньюман (1961), в живом организме встречаются те же три механизма перемещения ионов, которые характерны для оксиапатита in vitro:
- диффузия в гидратный слой, окружающий кристаллы;
- ионный обмен на поверхности кристаллов;
- внутрикристаллический обмен. Однако в отличие от условий in vitro в организме эти механизмы находятся под комплексным воздействием физиологических факторов — роста живой ткани, васкуляризации, перестройки и созревания кости, поступления минеральных веществ из кишечника, эндокринных механизмов.
Известно, что костные кристаллы отделены от внеклеточной жидкости несколькими фазами, между которыми обмен ионов происходит с разной скоростью. Внутренняя часть кристалла содержит ионы Са2+, РО43-, ОН— и, вероятно, небольшое количество других ионов, как Na+ или СО33-. Ионы здесь могут перемещаться только путем перестройки кристаллической сетки или рекристаллизации.
На поверхности кристалла находятся относительно посторонние ионы, удерживаемые в кристаллической решетке электростатическими силами поверхности (Na+, Sr2+, Co32-, F—, цитрат3-). Состав гидратационной оболочки, как водной фазы, окружающей кристаллы, отражает одновременно состав поверхности кристалла и внеклеточной жидкости. Последней фазой является внеклеточная жидкость органической матрицы. Наиболее быстро обмен происходит между внеклеточной жидкостью и гидратационной оболочкой, более медленно — между этой оболочкой и поверхностью кристалла, очень медленно — между поверхностью и внутренней частью кристалла.
Благодаря сорбционным механизмам и гетероионному обмену (например, Ca2+↔Na+) кость представляет собой важный резервуар свободных ионов. Она действует как буферная система в отношении таких электролитов, как Na+, Co32-, Mg2+, и участвует в гомеостатической регуляции Ca2+, РО43- и величины pH. В костной ткани содержится 98,5% всего кальция, 83% фосфора, до 70% магния и 40% натрия организма. Кроме того, в ней в том или ином количестве находятся более 30 микро- и ультрамикроэлементов. Одни из них входят в состав кристаллической решетки оксиапатита (фтор, цинк), другие (медь, молибден, кобальт) катализируют ферментативные реакции в остеобластах, третьи (марганец, ванадий и др.) принимают участие в энзиматических процессах минерализации коллагеновых фибрилл.
Показано, что процесс синтеза мукополисахаридов при физиологической регенерации сопровождается накоплением ряда микроэлементов (меди, марганца, кремния, алюминия, свинца), которые образуют хелатные комплексы с мукополисахаридами и, по-видимому, играют определенную роль в «созревании» коллагена и его дальнейшей пространственной агрегации. Вместе с тем многие остеотропные элементы, включая естественные радиоактивные элементы, по-видимому, являются: для скелета балластными.
В заключение следует сказать, что нерастворимые или труднорастворимые соединения, содержащие минеральные элементы, входят также в состав волоса, шерсти, перьевого покрова, роговых образований. Относительно форм этих соединений известно немного. Допускается миграция отдельных элементов в системе волос (перо) — кожа — кровь.