Жизнь на Земле имеет единое происхождение. Однако, говоря об этом как об одном из крупных обобщений биологии, нельзя забывать и о причинах ее разнообразия. Достаточно напомнить, что число видов на Земле ныне достигает порядка 2 млн., из них примерно 450 тыс. составляют растения. Следовательно, это разнообразие также нуждается в объяснении, исходя из принципа исторического развития живой природы.
Мы знаем, что растения и животные состоят из примерно одинаковых химических элементов, полимерных соединений (нуклеиновые кислоты, белки, липиды и др.) и имеют клеточную организацию; во многом сходно и строение их клеток. В то же время есть и существенные различия между растениями и животными, вытекающие из их образа жизни и организации. Достаточно запомнить о способности зеленых растений синтезировать органические вещества из неорганических. Существуют и другие различия — наличие жесткой оболочки у клеток, малая подвижность, широкое распространение вегетативного размножения у растений и т. п.
Значение растений для поддержания жизни на Земле чрезвычайно велико. Не было бы растений, вся нынешняя аэробная жизнь вымерла бы от нехватки пищи и кислорода. Так, в среднем на 1 м2 площади путем фиксации CO2 за год образуют органического вещества деревья около 1,4 кг, тропический лес — 2,3 кг, травы — 0,91 кг. Общий органический продукт растений в год на земной поверхности составляет 139 ∙ 109 т, на долю наземных растений приходится 110 ∙ 109, водных — 29 ∙ 109 т. При этом они выделяют более 115—250 млрд. т O2.
Сами растения неоднородны по организации, продолжительности жизни, темпам размножения, продуктивности и т. д. Несмотря на это, растения в целом выступают как средообразующий, фактор для других организмов. Поэтому в ходе эволюции живой природы растения оказывались пионерами в освоении суши и новых территорий. «Подтягивая» за собой представителей других царств на неосвоенные территории, растения, в свою очередь, получали мощный стимул для дальнейшего эволюционного развития. При этом складывались определенные взаимоотношения между разными организмами и формировались сами биогеоценозы как результат сопряженной эволюции видов (Г. А. Заварзин, 1993; В. К. Сергеев, 1993).
Способность к синтезу органических веществ из неорганических проявляется у разных живых существ, которые отличаются по организации и первичным механизмам процесса. Среди них встречаются одно- и многоклеточные, многоклеточные разной степени дифференциации; железобактерии, водородные бактерии и зеленые растения (водоросли, мхи, лишайники, папоротники, голосеменные и цветковые). Фотосинтез — одно из проявлений автотрофного обмена. По вопросу о возникновении автотрофного питания пока нет единого мнения, и здесь много неясного. Заманчива гипотеза К. С. Мережковского, А. С. Фаминцына, Б. М. Козо-Полянского и других о симбиогенезе и возникновении автотрофности путем симбиоза разных прокариот.
Показана общность молекулярных механизмов начальных стадий фотобиологических процессов у живых организмов (фотосинтез, фотоиндукционные и фоторегуляторные процессы). Во всех случаях имеют место поглощение света, миграция энергии, возбуждение фотоактивного хромофорного комплекса и т. д. (А. Б. Рубин, Г. Я. Фрайкин, 1987). Вообще фотоактивность заложена в природе самой жизни, а фотосинтез является лишь более специализированным ее проявлением, результатом действия отбора «на зачаточные стадии» полезных уклонений — фотоактивность. Последняя еще больше усиливается при наличии в клетке порфиринов (А. Лима де Фариа, 1991).
Одной из главных предпосылок перехода к автотрофному обмену была присущая живым организмам фотохимическая активность, обусловленная порфиринами. А. А. Красновский указывает, что даже бесхлорофилльные существа синтезируют фотохимически активные соединения — флавины и порфирины — как промежуточные продукты биосинтеза гема. Фотобиологические процессы характерны и для грибов. У них они протекают в том же спектральном диапазоне и с участием тех же веществ (флавины, металлопорфирины, тетрапироли), что и у зеленых растений (Г. Деборин и др., 1975).
Фотохимические реакции и возникающие свободные радикалы (С. П. Лоч, Б. Хейлс, 1979) первично могли участвовать в создании электрохимического потенциала ионов Н+ на поверхности мембран, энергия которого использовалась для синтеза АТФ и выведения избытка Ионов Na+ из клетки. В последующем указанный механизм дополняется «протонными насосами», что привело к усилению энергетических и транспортных возможностей клетки (Л. Н. Воробьев и др., 1980). Одна из гипотез возникновения протонных насосов исходит из возможности первичного появления пигмента в клетке и, следовательно, использования света при транспорте ионов Н+ через мембрану.
С появлением пигмента в клетке начинается автотрофная эра жизни, прошедшая через ряд ступеней совершенства. Напомним, что среди бактерий встречаются фотоорганотрофы (несерные пурпурные) и фототрофы (окрашенные серные бактерии, цианеи и др.), которые благодаря зеленому пигменту способны поглощать энергию и использовать ее для ассимиляции CO2. Фототрофные бактерии генетически неоднородны (ныне живущие фотосинтезирующие бактерии — обитатели водоемов). Поэтому полагают, что фототрофное питание первично возникло в водоемах (куда проникал свет) конвергентно в разных группах существ. Вероятно, по этой причине оно встречается у разнообразных групп прокариот, далеких в генетическом отношении. Из факта древности данного типа биоэнергетики вовсе не следует, что любая группа примитивных фотоавтотрофов должна быть признана филогенетически древней (М. В. Гусев, Г. Б. Гохлернер, 1980). Наиболее примитивные из них — пурпурные серобактерии — подвергают фотохимической ассимиляции ацетата без поглощения CO2:
Ацетат + КоА(Свет:NH3) Ацетил КоА → Органическая кислота → Аминокислота (глутамин)
Другим этапом автотрофного питания можно рассматривать способность восстанавливать CO2 при окислении органических продуктов, в частности изопропанола, пропанола, этанола, сахаров, органических кислот и т. п. (несерные пурпурные бактерии, серобактерии). В дальнейшем, по-видимому, возникла способность синтезировать органические вещества из неорганических (бактериальный фотосинтез), встречающаяся у многих пурпурных и зеленых серобактерий, которые окисляют сероводород, тиосульфат и т. д.
На стадии бактериального фотосинтеза фотоавтотрофный тип биоэнергетики заметно стабилизируется, что видно по соответствию величин квинтового выхода бактериального фотосинтеза и фотосинтеза зеленых растений (8—10 квантов на одну молекулу CO2).
Однако фоторедукция не связана с использованием воды в качестве донора электронов, так как у бактерий встречаются лишь фотосистема I (ФСI) и циклический транспорт электронов. Поглощенная энергия света идет у бактерий на синтез АТФ. Однако процессы фоторедукции имеют много общего с фотосинтезом, в том числе и восстановление CO2 при участии сильного редуцирующего фактора (Н), образовавшегося на свету. У бактерий нет ферментативных систем, способных выделять кислород в свободном виде из окисленных продуктов, и процесс идет так:
2OH + Н2А1 → 2Н2О + A1
Фоторедукция отличается от фотосинтеза еще и по донору водорода. Многие фоторедукторы в кислородной среде переходят к гетеротрофному типу питания, т. е. являются факультативными автотрофами. Фоторедукция предшествовала фотосинтезу, доказательством чего отчасти может служить способность ряда цианей, зеленых, красных и бурых водорослей переходить к фоторедукции в анаэробной среде. Переход от бактериального фотосинтеза к фотосинтезу водорослевого типа следует рассматривать как важный арахимоз в эволюции обмена. Это событие было вызвано диспропорцией между запасанием АТФ и восстановителя. Нехватка восстановителя оказалась фактором, ограничивающим бактериальный фотосинтез, и привела к разделению функций между пигментной и электрон-транспортной системами. Мутации, приведшие к разделению функций между указанными системами, могли возникнуть в популяциях подвижных фототрофов при перемещении их к поверхности водоема, где давление мутагенных факторов оказалось высоким (М. В. Гусев, Г. Б. Гохлернер, 1980).
Такой переход в биоэнергетике был сопряжен с дальнейшим повышением энергоемкости и конкурентоспособности организмов. Уже у синезеленых водорослей (цианей) формируются основные элементы аппарата фотосинтеза, переносчики энергии и ферменты. По некоторым данным, переход к фотосинтезу произошел еще раньше. Так, существуют химические доказательства фотосинтеза у организмов возраста 3,2 млрд. лет. Победе фотосинтетиков в борьбе за существование способствовало использование ими простыми легкодоступных веществ (CO2 и H2O) для создания органических молекул, синтез энергетически более богатых продуктов — углеводов в большем количестве, выделение свободного кислорода и т. д.
Несмотря на то, что цианобактерии (Cyanophyta) из-за приобретения способности к фотолизу воды достигают вершины биохимической эволюции фототрофных прокариот (М. В. Гусев, К. А. Никитина, 1979), они все еще сохраняют и другие метаболические механизмы: фиксацию азота, фотосинтез по бактериальному и растительному типу, ассимиляцию органических веществ (гетеротрофия) и хемосинтез. Например, у цианей (Anacystis nidulans и др.) при высокой интенсивности света, если отношение хлорофилла к фикоцианину составляет 1/3 CO2 фиксируется через цикл Кальвина, тогда как при слабом освещении или на красном свету — по С4-пути, а при концентрациях CO2 порядка 0,03% —через цикл карбоновых кислот. Тем не менее цианеи близки к зеленым растениям по составу пигментов, они восстанавливают НАДФ (вместо НАД у зеленых и пурпурных бактерий), имеют каротиноиды, пластоцианины, ферредоксины (близкие по составу к зеленым водорослям) и ФСI. Точно так же у многих водорослей в определенных условиях проявляются древние типы фотоавтотрофной ассимиляции: они способны переходить от фотосинтеза к фоторедукции (в атмосфере с водородом, азотом при низкой интенсивности света). Такое переключение типов питания часто сопровождается индукцией синтеза дополнительных ферментов и способствует повышению экологической пластичности вида.
Таковы вероятные этапы развития автотрофного питания. В ходе его эволюции на смену многообразным способам ассимиляции углерода приходит более специализированный способ — фотосинтез (филетическая эволюция). При этом на эффективность испытаны разные доноры электронов, источники энергии и структуры. В частности, Э. Либберт (1976) указывает, что эволюция фотосинтеза прошла три этапа: фотоорганотрофию (донор — органические вещества) → фотолитотрофию (донор — неорганические вещества) → фотогидротрофию (донор — вода). Эти ступени связаны с заметными сдвигами в кинетическом совершенстве организмов.
Таким образом, эволюцию автотрофного обмена можно представить так: гетеротрофы → облигатные фотогетеротрофы → факультативные фотогетеротрофы → облигатные фотоавтстрофы → фоторедукгоры → фотосинтетики.
Наиболее резкий скачок был совершен от сбраживателей к фотоорганотрофам. Между последними и фотолитотрофами границы считаются относительными.
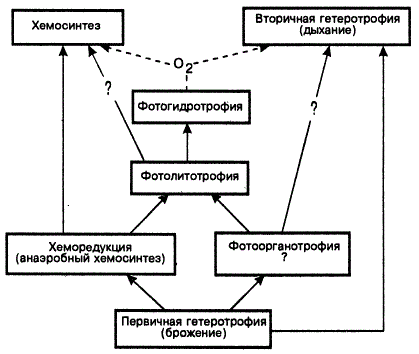
Эта схема неполно отражает связи, существующие между разными группами фототрофов. Эти этапы характеризуют экологическое совершенство, что, вероятно, и достигалось конвергентно у разных групп организмов. Эволюционно автотрофный обмен направлен на все большую специализацию. Напомним, что пурпурные бактерии наименее специализированы в отношении использования субстратов, тогда как у зеленых бактерий эти возможности ограничены. Как бы то ни было, в одной из групп прокариот (цианей) путем фотосинтеза достигнута полная независимость организмов от наличия готового органического вещества в среде.