Один из первых и наиболее ярких примеров взаимодействия генов был обнаружен в начале текущего столетия при анализе наследования формы гребня у кур.
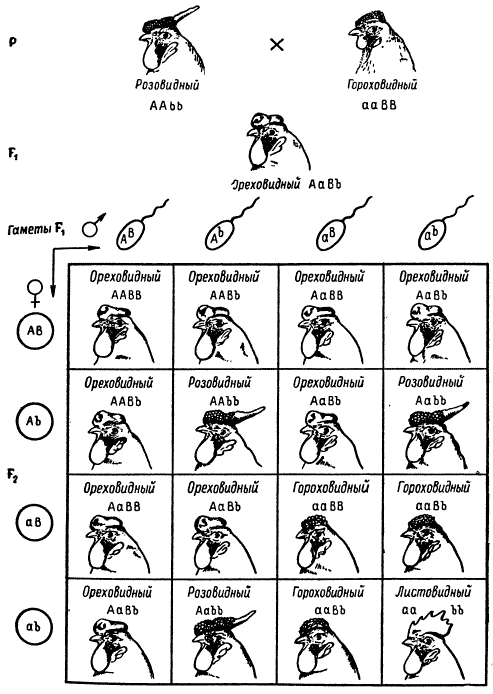
Каждая порода кур имеет определенное, характерное для нее, строение гребня. Так, например, леггорны обладают листовидным, или простым, гребнем, а белые виандотты имеют розовидный гребень — низкий, утолщенный спереди, заостренный сзади и у основания покрытый сосочками. Некоторые породы европейского прохождения обладают гороховидным гребнем — невысоким, с тремя продольными пластинками. У этих пород никогда не встречается называемый ореховидный гребень, напоминающий поверхность половинки грецкого ореха, характерный для кур малайского прохождения.
При скрещивании кур, имеющих розовидный гребень, с курами, имеющими простой гребень, в F1 доминирует розовидный тип. В F2 происходит расщепление в отношении 3 розовидных: 1 простой. Подобное же отношение получается и при скрещивании кур с гороховидным и простым гребнями. Доминантным признаком является здесь гороховидный тип. Следовательно, розовидный — простой гребень и гороховидный — простой гребень составляют две пары признаков, в которых простой гребень оказывается рецессивным. Когда скрещиваются куры, имеющие розовидный и гороховидный гребни, то их потомки в F1 оказываются с ореховидной формой гребня. Во втором поколении появляются куры с четырьмя типами гребней в отношении 9 ореховидных: 3 розовидных: 3 гороховидных: 1 листовидный, т. е. расщепление оказывается типичным для дигибридного скрещивания.
Наследование формы гребня у кур при взаимодействии двух генов
На основании этого следует предположить, что исходные породы с розовидным и гороховидным гребнями были гомозиготными и имели структуры ААbb и ааВВ, и в этом случае первое поколение должно быть гетерозиготным по этим двум генам, т. е. АаВb, которые при взаимодействии определяют развитие ореховидного гребня. Генетический анализ это полностью подтвердил.
Видно, что присутствие доминантных аллелей двух генов А и В у 9/16 кур второго поколения ведет к образованию ореховидного гребня. Присутствие гена А в гомо- или гетерозиготном состоянии (А—bb) определяет розовидную форму гребня у 3/16 особей, а присутствие другого гена В (ааВ—) обусловливает развитие гороховидной формы также у 3/16 особей. Гомозиготное состояние по обоим рецессивным генам aabb ведет к развитию простого листовидного гребня у 1/16 особей. Куры, имеющие этот гребень, при скрещивании друг с другом в последующих поколениях не дают расщепления. Зная закономерности наследования, определяющие форму гребня у кур, можно комбинировать гены путем скрещивания и получать желаемые типы гребней в определенном заранее рассчитанном отношении.
Поскольку в приведенном примере имеет место дигибридное расщепление (9:3:3:1), то это значит, что каждая пара аллелей ведет себя в наследовании независимо, и, следовательно, гены находятся в двух различных парах хромосом. Но действие доминантных или рецессивных аллелей этих двух генов, влияющих на проявление одного и того же признака, оказывается взаимосвязанным.
Взаимодействие генов является правилом, но оно мало изучено. В случае дигибридных скрещиваний расщепление в F2 по фенотипу может осуществляться в необычных отношениях: 9 : 7; 9 : 3 : 4; 13 : 3; 12 : 3 : 1; 15 : 1; 10 : 3 : 3; 9 : 6 : 1. Применение генетических и онтогенетических методов исследования дает возможность учесть различные типы взаимодействия генов и показать, что необычные расщепления по фенотипу в F2 представляют видоизменение все той же общей менделевской формулы 9:3:3:1.
Различают следующие основные типы взаимодействия генов: