Изучение бактерий открыло целый ряд явлений, осветивших с новой стороны источники наследственной изменчивости и механизмы наследственной передачи.
Одним из первых успехов в этой области было открытие явления трансформации у бактерий в 1928 г.
Известно несколько штаммов пневмококка Diplococcus pneumoniae: штамм S — с полисахаридной капсулой и гладкими колониями и штамм R — без капсулы и с шероховатыми колониями. Оба эти признака наследственны. Бактериолог Ф. Гриффитс инъецировал мышам вместе с убитым нагреванием штаммом пневмококка, обладающим капсулой (S), штамм живого пневмококка, лишенного капсулы (R). Спустя некоторое время ему удалось выделить из зараженных мышей живых пневмококков, обладающих капсулой. Таким образом, оказалось, что свойство убитого пневмококка — способность образовывать капсулу — перешло к живой бактерии. Поскольку признак наличия капсулы является наследственным, то следовало предположить, что какая-то часть наследственного вещества от бактерий штамма S перешла к клеткам штамма R. Но как это могло произойти, если клетки штамма S были убиты? Можно было предполагать, что в этом случае либо возникла мутация, либо произошла своеобразная гибридизация между живыми и мертвыми бактериями. Первое объяснение было наиболее вероятным, однако вопреки здравому смыслу второе объяснение оказалось ближе к истине.
В 1944 г. О. Эвери с сотрудниками удалось выяснить природу этого загадочного явления. Они взяли те же два штамма — R и S. Перед началом решающих опытов было изучено спонтанное мутирование обеих форм. Оказалось, что гладкая S-форма хотя и очень редко, но спонтанно мутирует в R-форму, а R-форма практически вовсе не мутирует в S-форму, т. е. мутации происходят почти исключительно в одном направлении: S→R. Но если R-форму помещали в экстракт из убитых клеток S-формы, то частота изменений R→S увеличивалась в 10 000 раз. Стало очевидным, что признак одного штамма (S) через какое-то вещество экстракта передавался другому штамму (R), т. е. возникало направленное наследственное изменение. Далее была произведена тщательная очистка — выделение этого вещества из экстракта клеток S-формы. Вещество было названо трансформирующим фактором (ТФ), а само явление — трансформацией.
Трансформирующий фактор по своей биохимической природе представлял собой не что иное, как дезоксирибонуклеиновую кислоту, входящую в состав хромосом. При этом было установлено, что он обладает некоторыми характерными свойствами. Его можно экстрагировать из клеток, очищать, воздействовать на него in vitro химическими и физическими факторами и затем снова вводить в живые клетки и изучать вызываемые им изменения.
Явление трансформации стало одним из основных доказательств роли ДНК как носителя наследственной информации. Теперь термином «трансформация» обозначают особый способ гибридизации бактерий, при котором происходит включение ДНК из клетки одного генотипа (донора) в клетку другого генотипа (реципиента), приводящий к рекомбинации генов. Иначе говоря, трансформация представляет включение вещества хромосомы донора в хромосому реципиента.
Сначала к этим исследованиям отнеслись скептически. Но вскоре многие исследователи поняли, что открыто не только новое явление, но и один из новых методов исследования наследственности. В последующих генетических и биохимических исследованиях было показано, что явление трансформации широко распространено у бактерий. Оно твердо установлено у самых различных видов и родов бактерий: Diplococcus, Staphyloccocus, Hemophilus, Neisseria, Agrobacterium, Rhizobium, Bacillus, Xantomonas.
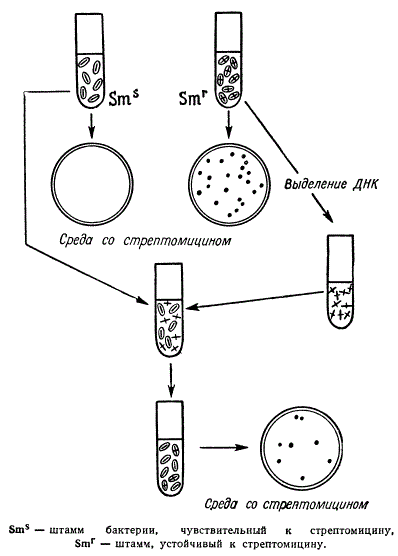
Схема опыта, демонстрирующего явление трансформации
Предположим, что один из штаммов пневмококков обладает наследственной чувствительностью к стрептомицину (реципиент). При рассеве чувствительных клеток на агаровой среде со стрептомицином колонии не развиваются. Другой штамм — Донор, устойчивый к стрептомицину, на среде с последним развивается. После раздельной инкубации этих штаммов из культуры донора выделяют ДНК, очищают ее от белка и вводят в культуру реципиента. Некоторые клетки реципиента оказываются восприимчивыми к ТФ и включают ДНК донора. Такие клетки называются компетентными клетками. После некоторого времени совместной инкубации чувствительных к стрептомицину пневмококков с ДНК Донора их высевают на агаровую среду со стрептомицином. При этом клетки реципиента, которые восприняли ТФ с геном устойчивости, дадут колонии. Клетки из этих колоний в дальнейшем бесконечно долго сохраняют устойчивость к стрептомицину. Эта устойчивость была передана с частью молекулы ДНК, где «записана» Наследственная информация стрептомициноустойчивости. Если же ДНК донора перед помещением в культуру реципиента обработать дезоксирибонуклеазой, то ТФ не передает наследственных свойств донора.
Активность трансформирующего фактора оказалась чрезвычайно высокой. Так, у Hemophilus трансформация осуществляется в течение 15 мин при концентрации ДНК 0,00015γ (γ = 10-6 г) в 1 мл среды. С помощью меченого фосфора (Р32) было показано, что не вся ДНК донора включается в геном реципиента, а лишь фрагменты с молекулярным весом около 3·105. В то же время под действием фермента дезоксирибонуклеазы (ДНК-азы), разрушающей ДНК, активность трансформирующего агента падает до нуля.
Трансформации могут подвергаться различные признаки. У пневмококков, например, трансформируется наличие капсулы, специфичность белков, размер и морфология колоний, устойчивость к антибиотикам (пенициллину и стрептомицину), способность к окислению определенных веществ и др.
Как правило, трансформируются отдельные свойства, но иногда одновременно несколько признаков в сцепленном состоянии. Р. Хочкис и Дж. Мармур при помощи ДНК, выделенной из штамма пневмококка, устойчивого к стрептомицину и способного сбраживать маннит, трансформировали оба эти свойства другому штамму пневмококка, не обладавшему ими. Одновременная передача обоих признаков от донора к реципиенту происходила в 50 раз чаще, чем это ожидалось, если бы трансформация по обоим признакам осуществлялась независимо. Дополнительная проверка показала, что действительно в рассматриваемом примере имеет место сцепленная передача обоих признаков.
Как правило, трансформация возможна между различными штаммами одного и того же вида, однако недавно была показана возможность межвидовой трансформации. В этом случае донором трансформирующего фактора были виды Hemophilus parainfluenzae или Н. aegypti, а реципиентом — Н. influenzae. Характерной особенностью межвидовой трансформации оказалась низкая частота ее осуществления в сравнении с внутривидовой.
При изучении действия мутагенов на ДНК, обладающую трансформирующей активностью, обнаружена различная чувствительность к мутагенам отдельных наследственных факторов этого трансформирующего фактора. Так, например, облучение ультрафиолетом значительно чаще инактивирует фактор, определяющий форму капсулы у пневмококков, чем фактор, обусловливающий устойчивость к стрептомицину.
Механизм трансформации еще недостаточно изучен. Предполагают, что при трансформации ДНК донора происходит как бы замена генов реципиента генами донора путем рекомбинации между молекулами ДНК. По аналогии с процессом рекомбинации у высших организмов в мейозе в отношении механизма трансформации предполагают, что трансформирующая ДНК, проникая в клетку-реципиент в виде осколков, конъюгирует с ее генетическим материалом. При этом одни авторы считают, что имеет место истинный обмен, сопровождающийся потерей одного из продуктов этого обмена. Согласно другой гипотезе «инфицирующий», или привнесенный, фрагмент ДНК непосредственно не включается в генетический материал, но направляет процесс репликации. При этом определенный участок ДНК клетки-реципиента воспроизводится по «матрице» ДНК донора.
Таким образом, трансформация обеспечивает генетическую рекомбинацию у бактерий. В этом может заключаться ее значение для эволюции бактериальных организмов. Обнаружение трансформации и изучение биохимической природы трансформирующего фактора явились вескими аргументами в пользу генетической роли ДНК как материального носителя наследственной информации.
После открытия явления трансформации у бактерий были сделаны попытки обнаружить это явление у высших животных. Получив экстракты ДНК из определенных тканей организма одного генотипа, их вводили другому в надежде на то, что специфическая ДНК донора вызовет в ДНК половых клеток реципиента направленное наследственное изменение. Хотя в этом плане было сделано несколько интересных попыток, убедительных фактов трансформации у высших организмов пока неизвестно. Впрочем, в принципе осуществление трансформации на соматических клетках животных и человека вполне возможно. Так, показано, что клетки в культуре тканей могут усваивать, включать меченую ДНК из среды. Возможно, метод культуры тканей откроет новые перспективы исследований в этой области.