До сих пор остается открытым вопрос о том, в какой момент митотического цикла происходит воспроизведение хромосом.
Как мы видели, даже в ранней профазе каждая хромосома оказывается удвоенной. Уже в середине профазы хромосомы состоят из двух хроматид. Это сестринские хроматиды — будущие хромосомы, но еще сдерживаемые единой центромерой. В прометафазе и метафазе каждая из хроматид оказывается также двойной. Эти половинки хроматид называют полухроматидами. Таким образом, в каждой хромосоме имеется не менее четырех хромонем.
Наиболее вероятно, что постройка хроматид и полухроматид осуществляется в интерфазе, и хромонемы, а может быть, и еще более тонкие структуры воспроизводятся в максимально деспирализованном состоянии. Следовательно, предполагается, что репродукция хромосомы осуществляется на уровне тонких нитей, а не целой хромосомы, как это считалось раньше.
В интерфазе каждая нить строит себе подобную. И если она была двойной, то каждая из таких двойных нитей оказывается учетверенной. Поэтому каждая состоит из жух хроматид, в каждой из которых имеется минимум по две полухроматиды, причем одна — от предыдущего деления ядра («старая» хроматида) и вторая — удвоенная в интерфазе данного митоза («молодая» хроматида). Так как число первичных хромонем в хромосоме в интерфазе всегда два или больше двух, то соотношение старых и молодых хромонем хромосомы останется одинаковым. Таким образом, в каждом митозе нет целиком заново построенных в данном цикле митоза дочерних хромосом. Каждая хромосома состоит из старых и молодых хромонем.
Удвоение хромосом рассматривают не только на хромосомном, но и на молекулярном уровне. Генетически и химически было установлено, что при удвоении хромосом содержание ДНК точно удваивается; это означает, что удвоение ДНК, или, как принято говорить, репликация ДНК, происходит в один из периодов цикла митоза. Хотя репликация ДНК в митозе не раскрывает полностью механизма удвоения хромосом, тем не менее этот процесс представляется главным в молекулярном механизме биосинтеза хромосом.
Изучение времени синтеза ДНК в клеточном цикле деления показало, что у многоклеточных организмов он происходит между двумя следующими друг за другом митотическими циклами. Цикл синтеза ДНК в клеточном делении, называемый генерационным циклом, приходится на период интерфазного состояния ядра.
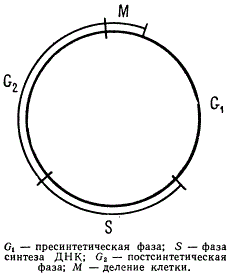
Соотношение цикла синтеза ДНК и клеточного деления
В первой фазе этого цикла, идущей вслед за прошедшим митозом и обозначаемой G1 не синтезируется ДНК, но осуществляется синтез РНК и белков. Затем следует фаза синтеза ДНК (фаза S), фаза, в течение которой количество ДНК в ядре клетки удваивается; потом наступает постсинтетическая фаза (фаза G2), когда ДНК не синтезируется, но идет синтез РНК и белков (в особенности ядерных) и накапливается энергия для следующего митоза. Этим цикл завершается и наступает профаза митоза.
Вводя в клетку (в определенный момент) предшественника ДНК — тимидин, меченный тритием, который в последующем включается в синтезируемую ДНК, по времени, прошедшем до появления первых меченых фигур митоза, устанавливают продолжительность всех фаз цикла биосинтеза ДНК. Время, в течение которого не появляются меченые фигуры микоза, соответствует фазе G2. Время, в течение которого начинают появляться такие митозы и до окончания возрастания их числа, соответствует периоду S, ибо только в фазе синтеза ДНК может происходить включение меченого тимидина. Иначе говоря, продолжительность фазы S определяется периодом, в течение которого увеличивается число треков радиоактивных меток на ядро. Зная продолжительность жизненного цикла данных клеток, можно высчитать продолжительность фаз.
Репродукция хромосом в митозе обеспечивает сходство вновь образованных клеток. Хромосомы являются пока единственными структурами, для которых доказана способность к строгому удвоению, поэтому генетика и рассматривает их как основу наследственности. Митохондрии, центросомы, пластиды и, видимо, другие органеллы клетки, также обладают свойством репродукции, так как и они содержат ДНК. Но их воспроизведение, очевидно, находится под контролем ядра.
Выше мы уже говорили, что абсолютное большинство исследователей склоняется к тому, что каждая хромосома сохраняет свою во всем клеточном цикле. Это представление подтверждается как морфологическими, так и генетическими факсами. Одним из прямых доказательств сохранения индивидуальности хромосом является то, что морфология каждой хромосомы и находящихся в ней генов сохраняется в непрерывном ряду митозов и смены поколений.
В пользу сохранения индивидуальности хромосом в интерфазе свидетельствует также следующий факт. Оказывается, что начинающаяся в телофазе деспирализация хромосом может происходить не полностью. Иногда после интерфазы, т. е. в профазе нового митоза, можно наблюдать не раскрутившиеся в телофазе предыдущего митоза спирали. Такие спирали называются остаточными (реликтовыми).
М. Дельбрук и Дж. Стент в 1957 г. предложили три схемы удвоения молекул ДНК:
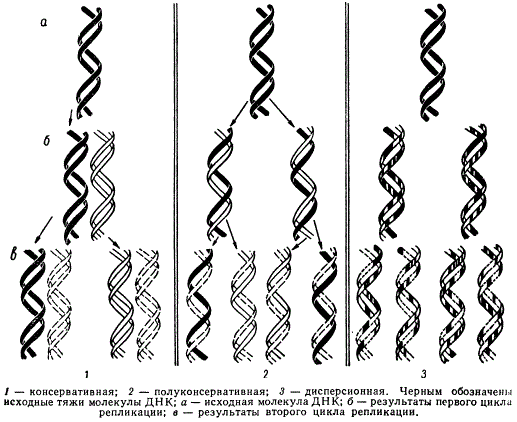
Возможные типы репликации (удвоения) ДНК
- консервативная схема — исходная двойная спираль ДНК остается неизменной и целостной в процессе синтеза и строит новую двухцепочечную молекулу;
- полуконсервативная схема — цепи двойной спирали молекулы ДНК расходятся, не разрываясь, и каждая из одиночных цепей ДНК служит матрицей для образования комплементарной цепи;
- дисперсионная схема — в процессе удвоения молекулы ДНК составляющие ее цепи разрываются или разрушаются, так что после синтеза дочерних молекул последние включают в свой состав случайным образом перекомбинированные фрагменты исходных молекул.
Ни одна из этих схем окончательно не доказана, однако полуконсервативная схема репликации ДНК представляется нам пока наиболее аргументированной, так как лучше других согласуется моделью структуры ДНК, разработанной Уотсоном и Криком. Согласно полуконсервативной схеме, при репликации ДНК сначала происходит разрыв водородных связей между пуриновыми и пиримидиновыми основаниями, образующими пары аденин-тимин и гуанин-цитозин. После разрыва двойная полинуклеотидная цепь раскручивается и каждая из образовавшихся одиночных строит около себя путем полимеризации комплементарную цепочку из нуклеотидов, находящихся в кариоплазме. В результате образуются две молекулы ДНК, идентичные исходной.
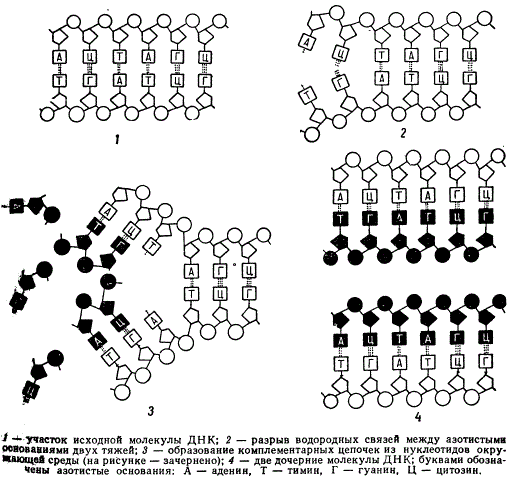
Схема, иллюстрирующая полуконсервативный механизм удвоения молекул ДНК
Таким образом, в самой двойственности структуры ДНК, комплементарности ее нуклеотидов заключено важнейшее условие репликации.
Изложенная гипотеза механизма репликации ДНК подтверждается опытами с применением изотопов. М. Мезельсон и Ф. Сталь разработали метод различения молекулы ДНК при включении в нее обычного азота N14 и более тяжелого его изотопа N15. Для этого бактерии Escherichia coli выращивают на источнике азота N15, а затем переносят на среду с N14. После ряда репликаций ДНК исследуют распределение двух составов ДНК по плотности с помощью центрифугирования в соответствующем растворителе. При этом тяжелая (с N15) и обычная (с N14) ДНК разделяются. По мере роста культуры происходит разделение полос ДНК с различной плотностью в градиенте плотности. Сначала появляется полоса, соответствующая изотопу N15, затем — промежуточная, соответствующая N15 и N14, и, наконец, соответствующая N14.
В этих опытах, а также в опытах на других объектах рядом авторов также было показано, что репликация ДНК происходит полуконсервативным способом.
М. Мезельсон и Ф. Сталь доказали также возможность гибридизации молекул ДНК, различающихся по указанным выше изотопам N14 и N15, т. е. с разным химическим составом. Для этого они взяли ДНК с промежуточной плотностью, т. е. случай, когда одна нить в двухнитчатой ДНК имела включенным N14, а другая N15, и с помощью нагревания разделили такую гибридную молекулу на две отдельные нити с разной плотностью.
Доказательства того, что митотическое деление ядра сопровождается репликацией ДНК, т. е. удвоением ее количества, и равным распределением между дочерними клетками, дали опыты Дж. Тейлора, где мечению (метод авторадиографии) подвергалась ДНК хромосом в период ее синтеза. В качестве специфической для ДНК метки был использован тимидин, меченный тритием. Этот метод позволил проследить судьбу исходных и дочерних хроматид в течение ряда последовательных митозов.
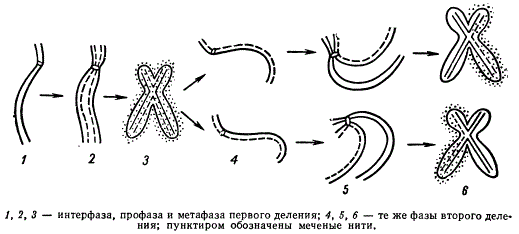
Схема редупликации хромосом, меченных тритием, в митозе (клетки корешков Vicia faba)
Сначала проростки семян конских бобов (Vicia faba) выращивали на среде, содержащей тимидин, меченный тритием. За это время хромосомы приобретали метку, которую можно видеть под микроскопом в виде зерен — треков на фотоэмульсии. Затем корешки бобов переносили в раствор колхицина, не содержащий радиоактивного тимидина. Колхицин обеспечивал сокращение хромосом до метафазного состояния и расчленение хроматид, тормозя расхождение дочерних хромосом. Просматривая препараты клеток корешков через определенные сроки после переноса их в раствор колхицина, определяли число происшедших удвоений хромосом в клетке. Зная, что у конских бобов диплоидное число хромосом равно 12, можно легко установить, что при наличии 24 хромосом удвоение их происходило 1 раз, а при 48 — 2 раза. При первом митозе, проходящем после включения метки» обе хроматиды каждой хромосомы оказались мечеными. Однако во втором митозе, протекавшем при отсутствии в среде меченого тимидина, только одна из двух хроматид каждой материнской хромосомы обнаруживала треки метки, поскольку включения метки во вновь редуплицированные хроматиды теперь уже происходить не могло. Эти исследования позволили сделать вывод о двойственном строении хромосом и о матричном механизме их воспроизведения.
Для объяснения механизма удвоения хромосом, как и для удвоения молекул ДНК, предложены три схемы:
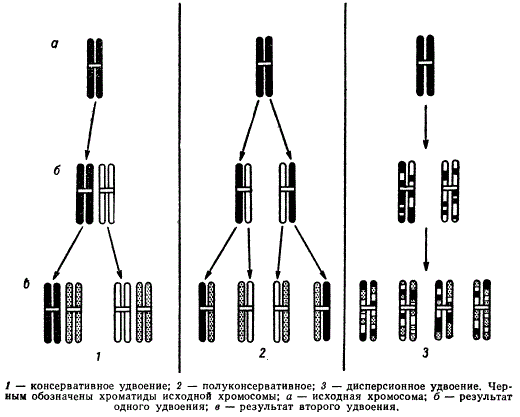
Возможные схемы удвоения хромосом
- консервативная схема — исходная двойная хромосома остается неизменной и целостной в процессе удвоения и строит новую двойную хромосому;
- полуконсервативная схема — половинки двойной хромосомы расходятся, не разрываясь, и каждая из них удваивается в процессе репродукции;
- дисперсионная схема — в процессе удвоения хромосомы составляющие ее половинки линейно распадаются на мелкие фрагменты, после удвоения которых дочерние хромосомы включают как новые, так и старые фрагменты.
В приведенных схемах хромосома рассматривается в момент, предшествующий ее редупликации, по крайней мере как двойная структура, так как во всех случаях хромосома содержит как минимум одну двуцепочечную молекулу ДНК.
Опыты Тейлора на Vicia faba позволяют считать наиболее вероятной полуконсервативную схему репродукции хромосом и что принцип полуконсервативной репликации ДНК, проверенный на молекулярном уровне, приложим и для воспроизведения хромосом. Данные Тейлора были подтверждены исследователями на хромосомах растений (Bellevalia, Crepis, Allium), животных (хомячка) и Человека. Схема полу консервативного удвоения хромосом хорошо согласуется со схемой полуконсервативной репликации молекулы ДНК, если допустить, что в момент, непосредственно предшествующий удвоению хромосомы, она состоит только из одной двухтяжевой молекулы ДНК. Но в настоящее время еще не ясно, как согласовать полуконсервативную схему удвоения хромосомы с полуконсервативной репликацией молекулы ДНК в том случае, если нередуплицированная хромосома состоит из многих двухтяжевых нитей ДНК.
Итак, хромосомы обладают некоторыми свойствами молекул: первичной химической структурой (расположением атомов и связями), вторичной линейной структурой, способной к спирализации, третичной — объемной (трехмерной) структурой. Хромосомы представляют собой сложные биополимеры. Они, по аналогии с молекулами, имеют определенный размер, относительное постоянство элементарного состава нуклеопротеидов и других компонентов, точную последовательность составляющих их частей, которые связаны в единую систему хромосомы.
Химический аппарат наследственности имеет молекулярную и субмолекулярную структуру. Признавая молекулярной основой репродукции хромосом репликацию ДНК, необходимо помнить, что самовоспроизведения (авторепродукции) хромосом нет, а есть сложный процесс их репродукции и репликации ДНК. Репликация ДНК осуществляется лишь при определенных условиях внутриклеточной среды. Поэтому связь здесь двусторонняя, а наследственная информация только в одном направлении — от гена к синтезируемой белковой молекуле.